Содержание
Введение………………………………………………………………………..…….2
Глава 1. Классификация синапсов и их характеристика …………………………3
1.1. Классификация синапсов…………………………………………………..
1.2. Структурно-функциональная характеристика синапсов………………….
Глава 2. Механизм синаптической передачи и ее регуляции ………..……5
Глава 3. Проведение возбуждения в синапсах ………………………….……….11
3.1. Проведение возбуждения в химических синапсах………………….……….11
3.2. Проведение возбуждения в электрических синапсах …………………….….1
Глава 4. Медиаторы и рецепторы синапсов ЦНС ……………….……………...15
Глава 5. Особенности физиологии синапсов у детей……………………………
Заключение……………………………………………………………………….…23
Список используемой литературы…………………………………………...……24
Введение
В нервной ткани имеются различные формы межклеточных контактов, среди которых главное место по функциональной значимости занимают синапсы. Основной функцией межклеточных контактов является их системообразующая роль, т.е. функция интеграции клеток в более сложные системы (тканевые, органные), что обеспечивается передачей от клетки к клетке различных сигналов (Семенов, 2005).
Синапс (греч. synapsis — соединение) — специализированная структура, обеспечивающая передачу возбуждающих или тормозных влияний между двумя возбудимыми клетками. Через синапс наряду с прямым влиянием на возбудимость иннервируемой клетки осуществляется и более медленное трофическое влияние, приводящее к изменению метаболизма иннервируемой клетки, ее структуры и функции. Понятие синапс как тип межклеточного соединения, при котором осуществляется перенос нервной информации, ввел в науку Ч. Шеррингтон (1897). По данным современной нейрофизиологии, в области синапсов происходят важнейшие процессы регуляции нейронной активности. Большое значение имеют синапсы в образовании условных связей, памяти, формировании пластичности нервных центров. Синапсы являются ареной деятельности многих лекарств, механизмов заболевания и выздоровления (Коробков, 1980).
Цель данной работы изучить механизмы передачи возбуждения в синапсах различных видов. Для достижения этой цели был поставлен ряд задач: рассмотреть структурно-функциональную характеристику синапсов, выявить роль медиаторов и рецепторов синапсов ЦНС.
Глава 1. Классификация синапсов и их характеристика
1.1. Классификация синапсов
Имеется несколько критериев, согласно которым классифицируются синапсы.
По виду соединяемых клеток синапсы можно разделить на межнейронные, нейроэффекторные и нейрорецепторные. Межнейронные синапсы находятся в ЦНС и вегетативных ганглиях. Нейроэффекторные (нейромышечные и ней-росекреторные) синапсы соединяют эфферентные нейроны соматической и вегетативной нервной системы с исполнительными клетками — поперечнополосатыми и гладкими миоцитами, секреторными клетками. К нейрорецепторным синапсам относят контакты во вторичных рецепторах между рецепторной клеткой и дендритом афферентного нейрона (Ашмарин,1999).
По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
По эффекту синапсы делят на возбуждающие, т.е. запускающие генерацию потенциала действия, и тормозные, препятствующие возникновению потенциала действия.
По способу передачи сигнала синапсы делят на химические, электрические и смешанные.
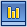
Химические синапсы являются специфическим межклеточным контактом для нервной системы. В них передача влияния на постсинаптическую клетку осуществляется с помощью химического посредника — медиатора. Этот тип синапсов преобладает в нервной системе человека и высших позвоночных, пример химического синапса проиллюстрирован на рисунке 1.
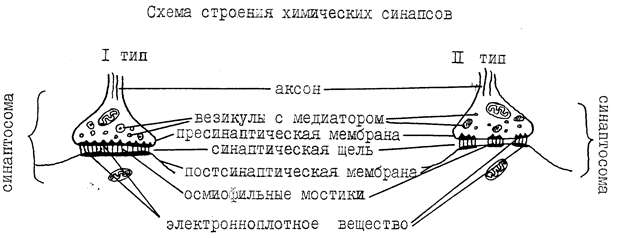
Рисунок 1. Строение химического синапса
( по: http://medlec.org/lek-68611.html)
В электрических синапсах потенциалы действия непосредственно (электротонически) передаются на постсинаптическую клетку. Эти синапсы являются разновидностью щелевых межклеточных контактов (высокопроводимые контакты), которые встречаются и в других тканях (например, нексусы в миокарде и гладкомышечной ткани). Электрические синапсы немногочисленны в нервной системе млекопитающих, особенно в постнатальном периоде. Пример строения электрического синапса представлен на рисунке 2.
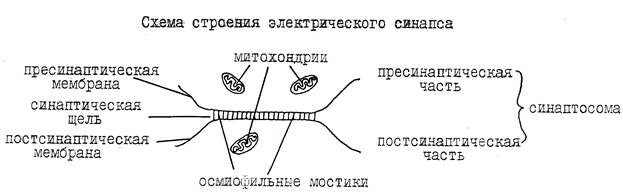
Рисунок 2. Строение электрического синапса
( по: http://medlec.org/lek-68611.html)
Обнаружены также смешанные синапсы, в которых наряду с химической передачей имеются участки с электротоническим механизмом передачи (например, в реснитчатом ганглии птиц, спинном мозге лягушки).
По природе медиатора химические синапсы делят на холинергические (медиатор — ацетилхолин), адренергические (норадреналин), дофаминергические (дофамин), ГАМКергические (у-аминомасляная кислота), глутаматергические (глутамат), аспартатергические (аспартат), пептидергичес-кие (пептиды), пуринергические (Волкова, 1983).
В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы различают: аксо-дендритические, аксо-соматические, аксо-аксональные, пример представлен на рисунке 3.
Рисунок 3. Классификация синапсов в зависимости от локализации окончаний терминальных веточек аксона.
( по: http://medlec.org/lek-68611.html)
1.2. Структурно-функцианальная характеристика синапсов
Нервно-мышечный синапс имеет общие для всех синапсов структурные элементы: пресинаптическое окончание, постсинаптическую мембрану и связывающую их синаптическую щель, пример строения синапса представлен на рисунке 4.
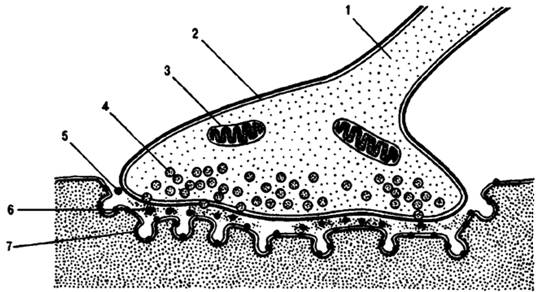
Рисунок 4. Нервно-мышечный синапс скелетной мышцы.
I — ветвь аксона; 2 — пресинаптическое окончание аксона; 3 — митохондрия; 4 — синоптические пузырьки, содержащие ацетилхолин; 5 — синаптическая щель; 6 — молекулы медиатора в синаптической щели; 7 — постсинаптическая мембрана мышечного волокна с рецепторами.
(по: Смирнову,2008г)
Вместе с тем структура нервно-мышечного синапса имеет и отличия от других синапсов, связанные с иннервацией длинных клеток (миоцитов) и необходимостью из одного синапса при передаче одного импульса практически одновременно активировать все сократительные единицы (саркомеры) миоцита (Россолимо, 2009).
Пресинаптическое окончание образуется расширениями по ходу разветвления аксона, иннервирующего мышечное волокно. В нервно-мышечном синапсе пресинаптическое окончание имеет большую длину (около 1—2 мм). Главным ультраструктурным фрагментом пресинаптического окончания являются синаптические пузырьки (везикулы) диаметром около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание и там заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 до 10 тыс. молекул химического вещества, участвующего в передаче влияния через синапс и в связи с этим названного медиатором (посредником). В нервно-мышечном синапсе везикулы преимущественно расположены вблизи периодических утолщений пресинаптической мембраны, называемых активными зонами. В неактивном синапсе везикулы с помощью белка синапсина связаны с белками цитоскелета, что обеспечивает их иммобилизацию и резервирование. Важными структурами пресинаптического окончания являются митохондрии, осуществляющие энергетическое обеспечение процесса синаптической передачи, цистерны гладкой эндо-плазматической сети, содержащие депонированный Са, а также микротрубочки и микрофиламенты, участвующие во внутриклеточном передвижении везикул. Часть мембраны пресинаптического окончания, ограничивающая синаптическую щель, называется пресинаптической мембраной. Через нее осуществляется выделение (экзоцитоз) медиатора в синаптическую щель (Губарь, 1993).
Синоптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость и мукополисахаридное плотное вещество в виде полосок, мостиков, которое обеспечивает связь между пре- и постсинаптической мембранами и может содержать ферменты. Это вещество хорошо выражено в щели нервно-мышечного синапса, где оно формирует базальную мембрану и содержит фермент ацетилхолинэстеразу.
Постсинаптическая мембрана — утолщенная часть клеточной мембраны иннервируемой клетки, содержащая белковые рецепторы, имеющие ионные каналы и способные связать молекулы медиатора. Ее особенностью в нервно-мышечном синапсе является наличие множества мелких складок, которые образуют слепые карманы, открывающиеся в синаптическую щель. Благодаря им резко увеличиваются площадь постсинаптической мембраны и количество ее рецепторов, которое в одном синапсе достигает 10—20 млн. Постсинаптическую мембрану нервно-мышечного синапса называют также концевой пластинкой (Самко, 2014).
Глава 3. Механизм синаптической передачи и ее регуляции
Передача в синапсе имеет два главных этапа.
Первый этап – это преобразование электрического сигнала в химический (электросекреторное сопряжение). Потенциал действия (ПД), поступивший в пресинаптическое окончание, вызывает деполяризацию его мембраны, открывающую потенциалзависимые Са-каналы. Ионы кальция входят, согласно концентрационному и электрическому градиентам, внутрь клетки, что ведет к увеличению его содержания в цитозоле в 10—100 раз. Ионы кальция активируют фосфорилирование синаптосина, что ослабляет связь везикулы с цитоскелетом, и везикула перемещается вдоль микротрубочек на позицию у активной зоны. При контакте везикулы с пресинаптической мембраной происходит ферментативное «плавление» ее стенки, а также активация белка синаптопорина, формирующего канал, через который медиатор выходит в синаптическую щель посредством первично-активного транспорта — экзоцитоза. В нервно-мышечном синапсе медиатором является ацетилхолин, который образуется в пресинаптическом окончании из ацетилкоэнзима А и холина под действием фермента холинаиетилтрансферазы. Впервые экспериментальное доказательство химического механизма передачи возбуждения в нервно-мышечном синапсе получил А.Ф.Самойлов (1924). Он показал, что скорость передачи возбуждения с нерва на мышцу в отличие от проведения возбуждения по нерву зависит от температуры в такой же степени, как и скорость химических реакций. Английский физиолог Г.Дейл (1934) установил, что медиатором нервно-мышечного синапса является ацетилхолин. Выделение молекул медиатора из пресинаптического окончания пропорционально количеству поступившего туда Са в степени п=4. Следовательно, химическое звено пресинаптического окончания работает как усилитель. Один из возможных механизмов усиления связан с тем, что поступивший в пресинаптическое окончание Са активирует рианодиновые рецепторы в цистернах эндоплазматической сети, имеюшие в своем составе Са-каналы, что приводит к дополнительному выделению Са в цитозоль из цистерн. Выделение ацетилхолина в синаптическую щель осуществляется квантами, каждый из которых в нервно-мышечном синапсе содержит от нескольких тысяч до 10 тыс. молекул. На один ПД из пресинаптического окончания нервно-мышечного синапса выделяется 200—300 квантов медиатора. В промежутках между ПД из пресинаптического окончания происходит спонтанное выделение 1—2 квантов медиатора в синаптическую щель в течение I с.
Молекулы медиатора, поступившие в синаптическую щель, диффундируют к постсинаптической мембране и вступают во взаимодействие с ее рецепторами. В нервно-мышечном синапсе ацетилхолин действует на Н-холинорецепторы, которые способны активизироваться и под влиянием никотина, вследствие чего они и получили свое название. Н-холинорецептор состоит из субъединиц и имеет в своем составе Na/K-канал. Скорость диффузии молекул медиатора позволяет им пройти расстояние синаптической щели в течение 0,1—0,2 мс. Длительность действия медиатора на рецепторы постсинаптической мембраны, определенная по продолжительности открытия в ней ионных каналов, равна около 1 мс. Это значительно меньше периода полураспада медиатора и свидетельствует о его удалении из синаптической щели. Оно осуществляется путем диффузии ацетилхолина из щели в окружающую жидкость и разрушения его под действием ацетилхолинэстеразы (Ашмарин,1999).
Этот фермент выделяется миоцитом и прикрепляется к мукополисахаридному веществу в синаптической щели. Одна молекула ацетилхолинэстеразы может гидролизовать до ацетата и холина 10 молекул ацетилхолина в 1 мс, что обеспечивает его разрушение в синаптической щели в течение нескольких десятых долей миллисекунды. При этом большая часть (около 60 %) холина захватывается обратно пресинаптическим окончанием (Сергеев ,1999).
Второй этап – это преобразование химического сигнала обратно в электрический. Этот этап осуществляется в постсинаптической мембране. Действие молекул медиатора на ее рецепторы ведет к открытию ионных каналов и перемещению ионов, имеющих высокий электрохимический градиент на протяжении канала. Присоединение двух молекул ацетилхолина к а-субъединицам Н-холинорецептора открывает канал. Открытое состояние сохраняется 1 мс, в течение которой через него проходит около 500 000 ионов. Канал на внутреннем суженном конце имеет диаметр 0,65 нм, хорошо проницаем для Na и К, плохо проницаем для Са2. Поскольку канал имеет слабую избирательность в отношении Na и К, то ионные токи через канал зависят главным образом от электродвижущей силы (ЭДС) этих ионов.
ЭДС иона равна разности между мембранным потенциалом покоя и равновесным потенциалом данного иона (ЭДС = МПП–Е ионна). Отрицательная величина ЭДС характеризует движение иона в клетку, положительная — из клетки.
В связи с этим входящий в клетку ток натрия (ЭДС = -140 мВ) резко преобладает над выходящим из клетки током калия (ЭДС = 14 мВ). Иными словами, ион Na движется в клетку согласно концентрационному и электрическому градиенту (клетка внутри имеет положительный заряд), а ион К выходит из клетки только согласно концентрационному градиенту, причем вопреки электрическому (снаружи клетка имеет положительный заряд). Поэтому суммарный ток ионов Na в клетку превосходит ток К из клетки, что и приводит к деполяризации постсинаптической мембраны. Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП), который в нервно-мышечном синапсе называют потенциалом концевой пластинки (ПКП), пример представлен на рисунке 5. Особенностью нервно-мышечного синапса фазного мышечного волокна является то, что при одиночной его активации формирующийся ПКП имеет большую амплитуду (30—40 мВ), которая превышает критический уровень деполяризации и вызывает генерацию ПД в миоците. Тоническое мышечное волокно имеет 7—10 синапсов, принадлежащих, как правило, нескольким мотонейронам. При этом ПКП не вызывает генерации ПД, а непосредственно запускает мышечное сокращение (Ткаченко, 1977).
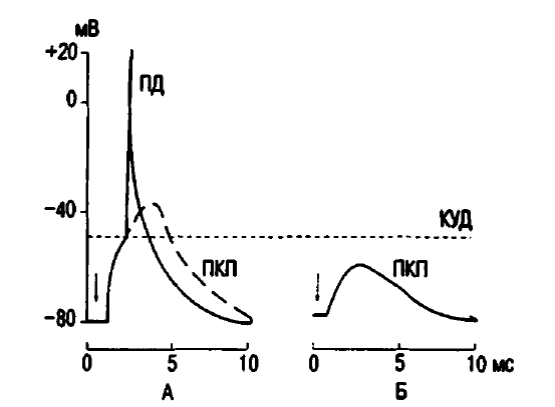
Рисунок 5. Потенциал концевой пластинки (ПКП). Стрелками показан момент нанесения стимула. КУД — критический уровень деполяризации: ПД — потенциал действия; А — ПКП в нормальной мышце, переходящий в ПД; Б — ослабленный ПКП, записанный в курарезированной мышце.
( по: Смирнову 2002 г.)
Как было отмечено, в промежутках между передачей нервного импульса происходит спонтанное выделение 1—2 квантов медиатора в синаптическую щель. При этом в постсинаптической мембране формируется деполяризация амплитудой 0,12—0,24 мВ, возникающая в среднем 1 раз в I с. Такие потенциалы, изученные в нервно-мышечном синапсе, были названы миниатюрными потенциалами концевой пластинки. Они, вероятно, поддерживают высокую возбудимость синапсов в условиях функционального покоя нервных центров. Кроме экзоцитоза медиатора, существует постоянная неквантовая утечка молекул медиатора в синаптическую щель. Предполагают, что неквантовая секреция играет трофическую роль.
Саморегуляция в синапсе осуществляется с использованием функциональных обратных связей. Веществами, влияющими на эффективность синаптической передачи, могут быть медиаторы, продукты их распада. В нервно-мышечном синапсе ацетилхолин, выделившийся в небольшом количестве в синаптическую щель, может стимулировать более сильный выброс ацетилхолина из пресинаптического окончания по механизму обратной связи (самоусиление секреции). Высокие концентрации ацетилхолина в синаптической щели, напротив, угнетают секрецию его из пресинаптического окончания (Караулова,2009).
Глава 3. Проведение возбуждения в синапсах
Рассмотрим проведение возбуждения в химических и электрических синапсах.
3.1 Проведение возбуждения в химических синапсах
Схема проведения возбуждения в химических синапсах представлена на рисунке 6.
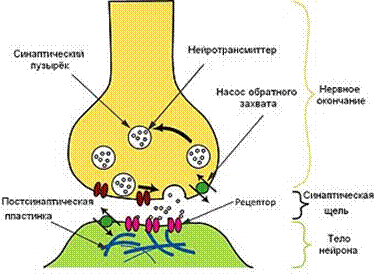
Рисунок 6. Схема проведения возбуждения в химических синапсах.
(по: http://medlec.org/lek-68611.html)
1. Синапсы функционально асимметричны и работают по принципу физиологического клапана, осуществляя одностороннее проведение возбуждения только в направлении от пресинаптического окончания в сторону постсинаптической мембраны. Это связано с тем, что медиатор выделяется из пресинаптического окончания, а взаимодействующие с ним рецепторы, имеющие ионные каналы, необходимые для формирования синаптических потенциалов, находятся только на постсинаптической мембране.
2. Имеется синоптическая задержка в проведении нервного импульса от нейрона на иннервируемую клетку, равная в нервно-мышечном синапсе 0,5—1,0 мс (время от момента поступления импульса к нервному окончанию до момента возникновения ПД в мышечном волокне). Это время затрачивается на процессы секреции медиатора, диффузию его к постсинаптической мембране, действие на рецепторы, возникновение ионных токов, формирование постсинаптических потенциалов и их суммацию, способную вызвать ПД.
3. Синапсы имеют низкую лабильность (посравнению с нервным волокном). Она равна около 100 Гц, что в 5—6 раз ниже лабильности аксона. Главной причиной низкой лабильности синапса является синаптическая задержка проведения возбуждения (Шульговский В.В., 1997).
4. В синапсах происходит трансформация ритма возбуждения: частота потенциалов действия, поступающих в синапс, обычно не совпадает с частотой ПД, генерируемых нейроном, имеющим данный синаптический вход. Однако в нервно-мышечном синапсе быстрого мышечного волокна трансформация ритма не выражена: один импульс нервного волокна вызывает один ПЛ в мышечном волокне.
5. Проводимость химических синапсов сильно изменяется под влиянием биологически активных веществ, лекарств и ядов. Например, в нервно-мышечном синапсе токсин возбудителя ботулизма подавляет синтез ацетилхолина в пресинаптическом окончании, угнетая обратное поглощение холина из синаптической щели. Кураре и курареподобные вещества (диплацин, тубокурарин и др.) обратимо связываются с Н-холинорецепторами постсинаптической мембраны, блокируя действие на нее ацетилхолина. Необратимо связывает холинорецепторы и полностью блокирует передачу возбуждения через синапс полипептид из яда змей а-бунгаротоксин. Ряд веществ (прозерин, фосфорорганические отравляющие вещества и инсектициды) подавляют активность ацетилхолинэстеразы, разрушающей ацетилхолин в синаптической щели. При небольшой ее инактивации происходят умеренное накопление ацетилхолина и облегчение синаптической передачи (например, действие лечебных доз прозерина). При большой инактивации анетилхолинэстеразы и значительном накоплении ацетилхолина синаптическая передача блокируется — развивается синаптическая депрессия ( Смирнов, 2010).
6. Синоптическое облегчение и депрессия. Эти изменения синаптической передачи возбуждения более детально изучены в нервно-мышечных синапсах, хотя имеют место и в синапсах ЦНС. Передача ПД через синапс, как было рассмотрено выше, сопровождается повышением концентрации Ca в пресинаптическом окончании, которая снижается до межимпульсного уровня в течение нескольких десятков миллисекунд. Если следующий ПД попадает в этот следовый период, то выброс медиатора в синаптическую щель увеличивается и формируется более высокоамплитудный ВПСП (в нервно-мышечном синапсе — ПКП), что приводит к повышению эффективности синаптической передачи — синоптическому облегчению).
Активация пресинаптического окончания может осуществляться ретроградными посредниками (окисью азота, арахидоновой кислотой, нейропептидами), которые выделяются постсинаптической клеткой. При передаче серии ПД через синапс концентрация Са оказывается повышенной и вблизи постсинаптической мембраны. Активация при этом Са зависимых ферментов (киназ, фосфатаз, протеаз) приводит к активации рецепторных белков и расщеплению белков, блокирующих рецепторы (например, белка фодрина, маскирующего глутаматные рецепторы). Облегчение синаптической передачи может быть связано также с увеличением синтеза рецепторов и, следовательно, их количества на постсинаптической мембране. Синаптическое облегчение является причиной оптимума частоты раздражения, открытого Н.Е.Введенским (1885) на нервно-мышечном препарате. В ЦНС синаптическое облегчение обозначается как феномен длительной потенциации. Он имеет важное значение в образовании условных рефлексов, формировании памяти и обучения (Коробков, 1980).
При более высоких частотах передачи импульсов через синапс (например, для нервно-мышечного синапса более 100 Гц) снижается эффективность синаптической передачи, что получило название •синоптическая депрессия» (пессимум Н.Е.Введенского) — блок проведения возбуждения в результате стойкой деполяризации постсинаптической мембраны мышечного волокна, поскольку механизмы инактивации ацетилхолина не успевают срабатывать (пессимальное торможение). Синаптическая депрессия может развиться и при редкой, но длительной активации синапса. Ее механизм на пресинаптическом уровне связывают с истощением запаса медиатора в пресинаптическом окончании, которого по расчетам хватает на 10 ООО синаптических передач и который может иссякнуть в течение нескольких минут. Другие механизмы депрессии связаны с накоплением высокой концентрации медиатора в синаптической щели вследствие того, что выброс медиатора в щель превышает возможности систем его разрушения и удаления. Высокий же уровень медиатора оказывает тормозящее влияние на секрецию его из пресинаптического окончания. Происходит также уменьшение чувствительности (дссенситизация) рецепторов постсинаптической мембраны к медиатору. Механизм десенситизации может быть связан с фосфорилированием рецепторов постсинаптической мембраны, что в несколько раз снижает их сродство к медиатору. Другим механизмом десенситизации является эндоцитоз комплекса медиатор+рецептор внутрь клетки. Поглощенные рецепторы могут опять встраиваться в мембрану (при ослаблении стимула) или разрушаться в лизосомах. Эти процессы затрудняют развитие ПД в постсинаптической клетке и, следовательно, могут привести к блокаде синаптической передачи (Башкиров ,1980).
Синаптическая депрессия является причиной пессимума частоты раздражения (торможение Введенского), открытого в исследованиях на нервно-мышечном препарате. В синапсах UHC она обозначается термином длительная депрессия и играет важную роль в торможении условных рефлексов и процессах забывания.
3.2. Проведение возбуждения в электрических синапсах
Электрические синапсы имеют щель на порядок меньше, чем химические синапсы; проводят сигнал в обе стороны без синаптической задержки; передача сигналов не блокируется при удалении Са; низкая чувствительность к фармакологическим препаратам и ядам; практическая неутомляемость ( как у нервного волокна). Очень низкое удельное сопротивление сближенных пресинаптической и постсинаптической мембранами обеспечивает хорошую электрическую проводимость за счет создания электрического поля. Определенный вклад в обеспечении хорошей электрической проводимости вносится коннексонами (Агаджанян,1998).
Глава 4. Медиаторы и рецепторы синапсов ЦНС
Медиаторами синапсов ЦНС являются многие химические вещества, разнородные в структурном отношении (в головном мозге обнаружено около 30 биологически активных веществ). Вещество, из которого синтезируется медиатор, попадает в нейрон или его окончание из крови или ликвора и в результате биохимических реакций под действием ферментов превращается в соответствующий медиатор, а затем транспортируется в синаптическую щел в виде пузырьков (везикул). Медиаторы синтезируются также в пресинаптических окончаниях. По химическому строению их можно разделить на несколько групп, главными из которых являются моноамины, аминокислоты и полипептиды. Достаточно широко распространенным медиатором в синапсах ЦНС является ацетилхолин (Белов ,2010)
Ацетилхолин встречается в различных отделах ЦНС ( кора большого мозга, спинной мозг). Известен в основном как возбуждающий медиатор. В частности, является медиатором а-мотонейронов спинного мозга, иннервирующих скелетную мускулатуру. С помощью ацетилхолина а-мотонейроны по коллатералям своих аксонов передают возбуждение на тормозные клетки Реншоу. В ретикулярной формации ствола мозга, в гипоталамусе обнаружены М- и К-холинорецепторы. При взаимодействии ацетилхолина с рецепторным белком последний изменяет свою конформацию, в результате чего открывается ионный канал. Тормозное влияние ацетилхолин оказывает с помощью М-холинорецепторов в глубоких слоях коры большого мозга, в стволе мозга, хвостатом ядре.
Амины ( норадреналин, дофамин, серотонин, гистамин) в большинстве своем в значитетельных колличествах содержатся в нейронах ствола мозга, в меньших количествах выявляются в других отделах ЦНС. Амины обеспечивают возникновение процессов возбуждения и торможения, например в промежуточном мозге, черной субстанции, лимбической системе, полосатом теле.
Норадреналин. Норадренергические нейроны сконцентрированы в основном в области голубого пятна ( средний мозг), где их насчитывается всего несколько сотен, но ответвления их аксонов встречаются по всей ЦНС. Норадреналин является тормозным медиатором клеток Пуркинье мозжечка и возбуждающим – в гипоталамусе, ядрах эпиталамуса.
Норадреналин регулирует настроение, эмоциональные реакции, обеспечивает поддержание бодрствования, учувствует в механизмах формирования некоторых фаз сна, сновидений (Ноздрачев,1991).
Дофамин учувствует в формировании чувства удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Дофамин полосатого тела регулирует сложные мышечные движения.
Серотонин. С помощью серотонин в нейронах ствола мозга передаются возбуждающие и тормозящие влияния, в коре мозга - тормозящие влияния. Имеется несколько типов серотониновых рецепторов. Серотонин реализует свое влияние с помощью ионотропных ( управляющих деятельностью ионных каналов) и метаботропных рецепторов, влияющих на биохимические процессы с помощью вторых посредников. Серотонин содержится главным образом в структурах, имеющих отношение к регуляции вегетативных функций.
Серотонин ускоряет процессы обучения, формирование болевых ощущений, сенсорное восприятие, засыпание; ангиотензин повышает артериальное давление, тормозит синтез катехоламинов, стимулирует секрецию гормонов; информирует ЦНС об осмотическом давлении крови.
Гистамин в довольно высокой концентрации обнаружен в гипофизе и срединном возвышении гипоталамуса - именно здесь сконцентрировано основное количество гистаминергических нейронов. Медиаторная роль изучена мало.
Аминокислоты. Кислые аминокислоты (глицин, гамма-аминомасляная кислота) являются тормозными медиаторами в синапсах ЦНС: глицин - в спинном мозге, ГАМК – в коре большого мозга, мозжечке, стволе мозга, спинном мозге. Нейтральные аминокислоты передают возбуждающие влияния и действуют на соответствующие возбуждающие рецепторы. Предполагают, что глутамат может быть медиатором афферентов в спинном мозге. Рецепторы глутаминовой и аспарагиновой аминокислот имеются на клетках спинного мозга, мозжечка, таламуса, гипокампа, коры большого мозга. Глутамат – основной возбуждающий медиатор ЦНС (75% возбуждающих синапсов мозга).
Полипептиды также выполняют медиаторную функцию в синапсах ЦНС. В частности, субстанция Р является медиатором нейронов, передающих сигналы боли. Особенно много этого полипептида содержится в дорсальных корешках спинного мозга. Это позволила предположить , что субстанция Р может быть медиатором чувствительных нервных клеток в области их переключения на вставочнные нейроны (Волкова,1983).
Энкефалины и эндорфины- медиаторы нейронов, блокирующих болевую импульсацию. Они реализуют свое влияние посредством соответствующих опиантных рецепторов, которые особенно плотно распологаются на клетках лимбической системы; много их также на клетках черной субстанции , ядрах промежуточного мозга и солитарного тракта, имеются они на клетках голубого пятна спинного мозга.
Эндорфины, энкефалины, пептид - дают антиболевые реакции, повышение устойчивости к стрессу, сон.
Ангиотензин учавствует в передаче информации о потребности организма в воде, люлиберин- в половой активности.
Олигопептиды – медиаторы настроения, полового поведения, передачи ноцицептивного возбуждения от переферии в ЦНС, формирование болевых ощущений.
Химические вещества, циркулирующие в крови – оказывают модулирующее влияние на активность синапсов. Простагландины, высвобождаемые из клеток, влияют на многие звенья синаптического процесса, например на секрецию медиатора, работу аденилатциклаз. Они обладают высокой физиологической активностью, но быстро инактивируются и поэтому действуют локально.
Гипоталамические нейрогормоны, регулирующие функцию гипофиза, также выполняют медиаторную роль (Данилова ,2009).
Принцип Дейла. Согласно принципу Дейла, каждый нейрон синтезируется и использует один и тот же медиатор или одни и те же медиаторы во всех разветвлениях своего аксона. Однако кроме основного медиатора в окончаниях аксона могут выделяться и другие, сопутствующие, медиаторы, играющие модулирующую роль и действующие более медленно. В спинном мозге установлено два быстродействующих медиатора в одном тормозном нейроне – ГАМК и глицин, а также один тормозной ( ГАМК) и один возбуждающий ( АТФ). Принцип Дэйла звучит так: « один нейрон – один быстрый медиатор».
Эффект действия медиатора зависит в основном от свойств ионных каналов постсинаптической мембраны и вторых посредников. Это явления особенно ярко демонстрируется при сравнении эффектов отдельных медиаторов в ЦНС и периферических синапсах организма. Ацетилхолоин, например, в коре головного мозга при микроаппликациях на разные нейроны может вызывать возбуждение и торможение, в синапсах сердца – торможение, в синапсах гладкой мускулотуры желудочно-кишечного тракта – возбуждение. Катехоламины стимулируют сердечную деятельность, но тормозят сокращение желудка и кишечника (Сергеев ,1999).
Глава 5. Особенности физиологии синапсов у детей
Структурно-функциональное созревание нервно-мышечных синапсов охватывает период антенатального и раннего постнатального периодов развития. Оно имеет свои особенности для пресимпатической и постсинаптической мембран.
А. Созревание пресинаптической мембраны. Двигательные нервные окончания в мышцах появляются на 13—14-й неделе внутриутробного развития. Формирование их продолжается длительное время и после рождения. Мышечное волокно новорожденного, как и взрослого человека, имеет один синапс в виде типичной концевой бляшки. Созревание пресинаптической мембраны проявляется в увеличении терминального разветвления аксона, усложнении его формы, увеличении площади всего окончания. Степень созревания нервных окончаний значительно увеличиваются к 7—8 годам, при этом проявляются более быстрые и разнообразные движения.
В процессе развития усиливается синтез ацетилхолина в мотонейронах, увеличиваются количество активных зон в пресинаптическом окончании и количество квантов медиатора, выделяющегося в синаптическую щель.
Б. Созревание постсинаптической мембраны. Когда окончание аксона достигает миотрубки, в соответствующем ее участке появляются скопления митохондрий, рибосом, микротрубок. На поверхности миотрубки образуются выемки, в которых помещается окончание аксона. Образуется примитивное нерв-но-мышечное соединение. Особенностью ранних стадий развития мышечных волокон является разлитая чувствительность всей поверхности мембраны к ацетилхолину, присущая донервной стадии развития мышц. В процессе развития на каждом мионе сохраняется единственный синапс, формирование его сопровождается появлением в постсинаптической мембране холинэстеразы, увеличение концентрации которой приводит к повышению скорости гидролиза ацетилхолина. Образуются складки на постсинаптической мембране, в результате чего растет амплитуда ПКП и повышается надежность передачи возбуждения через синапс. Внесинаптическая поверхность мышечного волокна постепенно теряет чувствительность к ацетилхолину. После денервации в эксперименте чувствительность к ацетилхолину вновь распространяется на всю поверхность мембраны (Батуев ,2009).
Вследствие незрелости нервно-мышечного синапса у плода и новорожденного синаптическая передача возбуждения происходит медленно. Без трансформации ритма через такой синапс передается не более 20 импульсов в 1 с, а к 7—8 годам жизни — около 100 импульсов в I с, т.е. как у взрослого.
Утомляемость синапса объясняется истощением медиатора и снижением чувствительности постсинаптической мембраны к медиатору в результате накопления продуктов обмена, закислением среды — все это приводит к снижению ПКП (Башкиров ,1980).
Заключение
В работе была рассмотрена структурно-функциональная характеристика синапсов.
Показано, что синапсы классифицируются: по виду соединяемых клеток, по эффекту, по способу передачи сигналов, в зависимости от местоположения в ЦНС.
Были изучены механизмы передачи возбуждения в синапсах различных видов, в частности химических и электрических.
Установлено, что химические синапсы характеризуются односторонним проведением возбуждения, замедленным распространением возбуждения в ЦНС, низкой лабильностью, утомляемостью и их действие легко блокируется фармакологическими препаратами.
В свою очередь, электрические синапсы проводят сигнал в обе стороны без синаптической задержки, имеют низкую чувствительность к фармакологическим препаратам, обладают практической неутомляемостью.
В работе были приведены характеристики медиаторов и рецепторов синапсов ЦНС, их роль в организме.
Список используемой литературы
1. Ашмарин, И.П. Биохимия мозга: Учеб. Пособие / И. П.Ашмарин, П.В. Стукалова, Н.Д. Ещенко. – СПб., 1999. – 432с.
2. Курепина, М.М. Анатомия человека: Учебник для биологических факультетов педагогических институтов / М.М. Курепина, Г.Г. Воккен. – М.: Просвещение, 1979. – 304с.
3. Волкова, О.В. Эмбриональный гистогенез и постнатальное развитие органов человека / О.В. Волкова. – М., 1983. – 123с.
4. Сергеев, П.В. Рецепторы физиологически активных веществ / П.В. Сергеев, Н.Л. Шимановский, В.И. Петров. – М.; 1999.
5. Бабский, Е.Б. Физиология человека: учебник для медицинских институтов / Е.Б. Бабский, В.Д. Глебовский, А.Б. Коган. – Москва: Альянс, 2015. – 560 с.
6. Самко, Ю.Н. Физиология [Электронный ресурс]: Учебное пособие / Ю.Н. Самко. – М.: НИЦ ИНФРА-М, 2014. – 144 с.
7. Ткаченко, Б.И. Основы физиологии человека / Б.И. Ткаченко – СПб.,1994. 114с.
8. Батуев, А. С. Физиология высшей нервной деятельности и сенсорных систем: учебник / А. С. Батуев. - СПб.: Питер, 2009. - 317 с.
9. Данилова, Н. Н. Психофизиология: Учебник / Н. Н. Данилова. - М. : Аспект Пресс, 2000. - 373с.
10. Караулова, Л. К. Физиология : учебное пособие / Л. К. Караулова, Н. А. Красноперова, М. М. Расулов. - М. : Академия, 2009. - 384 с.
11. Семенов, Э. В. Физиология и анатомия : учебное пособие / Э. В. Семенов. - М.: Джангар, 2005. - 480 с.
12. Смирнов, В. М. Физиология центральной нервной системы: учебное пособие / В. М. Смирнов, В. Н. Яковлев. - М. : Академия, 2002. - 352с.
13. Смирнов, В. М. Физиология человека: учебник / В. М. Смирнова. - М. : Медицина, 2002. - 608с.
14. Россолимо, Т. Е. Физиология высшей нервной деятельности : хрестоматия: учебное пособие / Т. Е. Россолимо, И. А. Москвина - Тарханова, Л. Б. Рыбалов. - М.; Воронеж: МПСИ: МОДЭК, 2007. - 336 с.
15. Белов, А.Ф., ред. Системный подход в изучении интегративной деятельности мозга научн. труды / А.Ф. Белов. РМИ. - Т.71. — Рязань, 1980. - 98 с.
16. Ноздрачев, А.Д. . Общий курс физиологии человека и животных: учебник / А.Д. Ноздрачев. — М.: Высшая школа, 1991, т. 1, 528 е.; т. 2, 528 с.
 (zip - application/zip)
(zip - application/zip)