Министерство образования и науки РФ
Федеральное государственное автономное образовательное учреждение высшего образования
«КАЗАНСКИЙ (ПРИВОЛЖСКИЙ) ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ»
ИНСТИТУТ ФУНДАМЕНТАЛЬНОЙ МЕДИЦИНЫ И БИОЛОГИИ
КАФЕДРА БОТАНИКИ И ФИЗИОЛОГИИ РАСТЕНИЙ
Направление: 06.03.01 – биология
КУРСОВАЯ РАБОТА
ИВАНОВОЙ АИДЫ АНАТОЛЬЕВНЫ
Видовое разнообразие макромицетов Раифского Волжско – Камского государственного природного биосферного заповедника
Студент 3 курса
Группа 01-206
"31" октября 2015 г. ____________________ (А.А. Иванова)
Научный руководитель
"31" октября 2015 г. ____________________ (К.О. Потапов)
Казань – 2015
СОДЕРЖАНИЕ
Введение. 3
Глава 1. Обзор литературы.. 6
1.1. Характеристика анатомического, морфологического строения афиллофороидных грибов. 6
1.2. Характеристика анатомического, морфологического строения агарикоидных грибов. 8
1.3. Характеристика анатомического, морфологического строения гастероидных грибов 10
1.4. История изучения грибов в Республике Татарстан. 12
1.5. Географическая характеристика ВКГПБЗ. 16
Глава 2. Материалы и методы.. 19
Глава 3. Результаты исследования. 25
3.1. Систематическая структура. 25
3.2. Трофическая структура. 34
Выводы.. 39
Список используемых источников. 40
Введение
Одним из важных свойств жизни является ее разнообразие. Так, среди многих проблем современной биологии изучение и охрана биоразнообразия планеты является одной из самых важных. При этом наиболее актуальными задачами микологии по-прежнему является инвентаризация видового состава микобиоты, выявление закономерностей географического распространения грибов и их экологических особенностей.
Предполагаемое разнообразие видов грибов достигает 1,5 млн., из которых описано лишь около 5% [Черепанова, 2005]. Не малую долю макромицетов составляют агарикоидные, афиллофороидные и гастероидные базидиомицеты. Эта группа грибов включает около 30 тысяч видов, разнообразных по морфологическим, анатомическим признакам и экологическим предпочтениям [Малышева, Малышева, 2008]. Известно примерно такое же количество видов аскомицетов, к группе которой относятся дрожжи, сморчки, трюфели и строчки.
Грибы, как гетеротрофные организмы, принимают активное участие в лесном круговороте веществ. К ним относятся микоризообразующие, паразитические и сапротрофные виды, каждая группа из которых занимает свое особое положение в экосистеме. Грибы-микоризообразователи, находятся в симбиозе с корнями микотрофных древесных пород, к которым относятся все лесообразующие породы Раифского леса, а именно сосна, ель, береза, дуб и липа. Не маловажную роль играют грибы в жизни своих симбионтов, которые получая углерод снабжают деревья необходимыми элементами, такими как азот, фосфор, кальций, магний, калий и другими катионами. Важное значение в жизни лесных фитоценозов выполняют подстилочные сапротрофы разлагающие опад, падающий на землю, в среднем, по несколько тонн на гектар. Грибы - микоризообразователи, гумусовые и подстилочные сапротрофы имеют положительное значение в жизни леса, что нельзя сказать о паразитических грибах. Представители этой группы могут относиться, как к полезным, так и к вредным, которые попадая через рану на обнажившуюся древесину губят дерево [Васильева, 1977]. В настоящее время агарикоидные и афиллофороидные базидиомицеты считаются важным компонентом экосистемы, которые выполняют главную роль в разложении и гумификации древесины, опада и остатков высших растений [Малышева, Малышева, 2008].
Таким образом, значение грибов в природе и их высокое разнообразие рождают высокую потребность в детальном изучении их экологии и географии. Однако на территории России подобные исследования проводились крайне неравномерно. Что касается территории Среднего Поволжья, то здесь грибы планомерно изучались в большей части областей и республик, за исключением Чувашии и Мордовии. К примеру, в 2008 - 2013 гг. на территории Удмуртии впервые для республики были отмечены 240 видов макромицетов. Целью последних исследований в Удмуртской Республике является инвентаризация ее микобиоты. В связи с чем, выявлено около 600 видов сумчатых и базидиальных макромицетов [Капитонов, 2013]. Что касается Кировской области, то сведения по афиллофороидным базидиомицетам приводятся в статье И. В. Ставишенко и Е. А. Лугининой «Афиллофороидные грибы государственного природного заказника «Былина» », где опубликованы первые сведения о видовом составе афиллофороидных грибов заказника. К настоящему времени выявлено 175 видов этой группы, из которых 58 видов впервые встречаются в Кировской области [Ставищенко, Лугинина, 2015]. В 214 году, в честь трехсотлетия Ботанического института им. В. Л. Комарова РАН издается монография, которая открывает серию публикаций, посвящённых агарикомицетам Приволжской возвышенности. Почти не исследованной остается Республика Мари Эл.
В Республике Татарстан, не смотря на довольно богатую историю изучения грибов, по-прежнему остается актуальным исследование биоразнообразия, поскольку большая часть работ на эту тему носит локальный характер, зачастую ограничена либо отдельным семейством или порядком, либо незначительной по площади территорией. Таким образом, современные исследования позволяют выявлять все новые и новые виды для микобиоты республики, ранее не указывающиеся для ее территории.
Цель настоящей работы: изучение видового состава макромицетов в лесных экосистемах ВКГПБЗ.
В связи с этим, были поставлены следующие задачи:
1. произвести сбор и идентификацию макромицетов;
2. определить особенности таксономической структуры биоты высших базидиомицетов на исследуемой территории;
3. проанализировать эколого-трофическую структуру выявленной микобиоты;
4. выявить редкие и новые для обследуемой территории виды грибов-макромицетов.
Проведение исследований в данном направлении позволит выявить закономерности формирования микобиоты высших базидиомицетов, в экологическом и географическом аспектах.
Глава 1. Обзор литературы
1.1. Характеристика анатомического, морфологического строения афиллофороидных грибов
В лесных экосистемах афиллофороидные грибы играют доминирующую роль в разложении древесины. Некоторые представители этого класса имеют способность к образованию эктомикориз [Змитрович, 1999].
Группу высших базидиальных грибов называют афиллофороидными макромицетами, представители которой характеризуются наличием базидиом с гладким, шиповидным, зубчатым, складчатым, пластинчатым или же трубчатым гименофором. Многие афиллофороидные грибы характеризуются своей способностью к многолетнему существованию, за счет дифференциации гиф плодового тела [Спирин, 2003]. Данная группа грибов очень велика и разнообразна. Она включает в себя виды с многоклеточным мицелием, который часто видоизменяется. Эти изменения помогают грибам в неблагоприятных условиях и в период размножения. В определенный период развиваются споры, которые образуются в плодовых телах грибницы [Клюшников, 1957].
Плодовые тела трутовиков очень разнообразны. Распростертые, распростерто-отогнутые или сидячие - в каждой из данных форм существуют свои модификации. Распростертые базидиомы бывают от тонких до почти подушковидных, с еле заметной или же толстой подстилкой. Сидячие или распростерто-отогнутые базидиомы могут развиваться как одиночно, так и группами. Размер распростертых базидиом варьирует от миллиметров до десятков сантиметров [Бондарцева, Пармасто, 1986; Бондарцева, 1998].
Важнейшими признаками нересупинатных плодовых тел является поверхность шляпки, она может быть голой, гладкой, шероховатой или бархатистой. Так же, не менее важна окраска. Ее пределы от блеклых светлых тонов, до разнообразных ярких цветов.
Важным признаком на уровне рода является консистенция и окраска ткани. Она может быть желатиновой, мясистой, пробковидной и др. Бывают случаи, когда наблюдается двухслойная ткань. В системах прошлого времени главным таксономическим признаком являлась конфигурация гименофора, так называемой части базидиомы, где развивается гимениальный слой. Основная форма гименофора, у грибов трутовых семейств, трубчатая.
Признак гифальной системы определяется степенью дифференциации гиф в базидиоме. Самым простым исходным типом называют мономитическим, а гифы составляющие базидиому - генеративными. Димитическое строение характеризуется сочетанием двух типов гиф: генеративно-скелетных и генеративно-связывающих. В зрелой стадии генеративные базидиомы могут быть обнаружены как обрывок на концах скелетных гиф в растущем крае, так как связывающие и скелетные гифы полностью вытесняют их [Бондарцева, 1961; Бондарцева, 1998].
Одним из самых важных таксономических признаков является строение спор. Они могут различаться по форме, размерам, окраске, орнаментации или по толщине оболочки [Бондарцева, 1998].
И так, у афиллофороидных грибов есть ряд микро- и макропризнаков характеризующие их родство.
1.2. Характеристика анатомического, морфологического строения агарикоидных грибов.
В
отличие от афиллофороидных грибов, базидиомы у агарикоидных грибов менее
разнообразны (рис. 1, [http://hrib.ru/Bazidioma/id/104-Vneshniy-vid-plodovyih-tel.html])
. Составными частями типичной базидиомы являются ножка, шляпка, а так же
расходящиеся мясистые пластинки или же плотно прижатые друг к другу трубочки [Мюллер,
Лёффлер, 1995].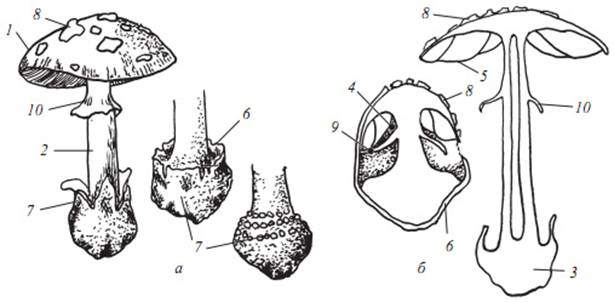
Рисунок
1. Строение базидиом мухоморовых грибов:
а - общий вид базидиомы; б - разрез молодого и зрелого плодового тела; 1 - шляпка;
2 - ножка; 3 - основание ножки; 4 - пластиночная камера; 5 - пластинчатый
гименофор; 6 - общее покрывало; 7 - вольва; 8 - остатки общего покрывала в виде
чешуй на шляпке; 9 - частное покрывало;10 - кольцо на ножке, остатки частного
покрывала.
Шляпки грибов может быть слизистой, сухой, гладкой или войлочной. Окраска всего плодового тела может быть очень разнообразна, в зависимости от содержащихся пигментов. Диаметр варьирует от нескольких миллиметров до десятки сантиметров. Край шляпки бывает ребристым, гладким, с остатками частного покрывала.
Характеристика гименофора является важным систематическим признаком. В определённом гименофоре пластинки могут быть толстыми или тонкими, края ровные, волнистые или зубчатые. Тип прикрепления пластинок может быть свободным, слабо и сильно низбегающие, прикрепленные зубцом и т.д. Бывает, пластинки срастаются между собой, образовывая кольцо вокруг ножки - коллариум.
Ножка, как и шляпка, бывает различной формы. Может расширяться или сужаться к основанию, а у некоторых древесных агариковых грибов может быть еле заметна или отсутствовать вообще [Переведенцева, 2007].
К главным микроскопическим признакам относят цистиды в гимениальном слое, размеры и формы спор, пряжки на гифах или их отсутствие, строение пилеипеллиса и стипитипеллиса, форма базидий и число стеригм на них [Нейздойминого, 1996; Переведенцева, 2007].
1.3. Характеристика анатомического, морфологического строения гастероидных грибов
Гастеромицеты - это группа грибов, чей мицелий состоит из сплетения разветвленных гиф, пронизывающих субстрат (рис. 2, [http://uvelicheniesuper.zapto.org/twins/?index=phrc&tht]). Гифы, обычно, белого цвета и не имеют специальных защитных приспособлений. Их форма приближена к округлой: грушевидная, шаровидная или же имеется ножка с шляпкой или булавовидным утолщением. Перидий [покров] гастероидных грибов бывает простым, двойным или многослойным. Высокоорганизованные наземные гастеромицеты имеют дифференцированный двойной перидий, состоящий из экзоперидия [внешний слой] и эндоперидия [внутренний слой]. Поверхность внешнего слоя может быть гладкой, чешуйчатой, шиповидной или бородавчатой. Ножка может, как присутствовать, так и отсутствовать или же быть просто результатом срастания ризоморф [Шварцман, Филлимонова, 1970; Сосин, 1973].
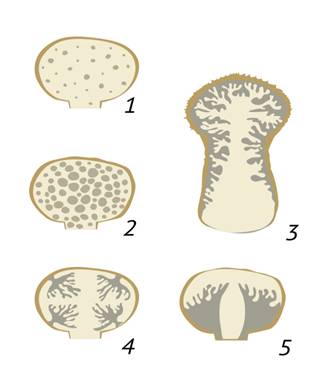
Рисунок 2 Типы плодовых тел гастеромицетов. 1. Равномерный; 2. Лакунарный или камеровидный; 3. Коралловидный; 4. Многошляпочный; 5.Одношляпочный.
Глеба - содержимое плодового тела, которое участвует в спорообразовании. С течением времени она может разрушаться, образуя споры, но зачастую остается в виде гиф и образует так называемый капиллиций, который бывает разветвленным, неразветвленным или представляет собой сеть с утончающимися гифами различного порядка. У гастероидных грибов с лакунарным типом строения плодового тела, иногда, имеется стерильный вырост внутри базидиомы - колонку или колумеллу. Ее форма и длина являются важным признаком в определении вида и рода грибов данной группы.
Гимений гастероидных грибов выстилает поверхность лакун, имеет базидии различной формы, зачастую короткобулавовидные. У редких видов присутствуют цистиды. Споры, часто, орнаментированы, но встречаются и гладкие [Ребриев, 2002].
1.4. История изучения грибов в Республике Татарстан
Полтора века на территории Республики Татарстан проводились исследования, которые связаны с изучением биоразнообразия грибов, их природном значении и искусственных экосистемах. С именами Н. В. Сорокина, П. Н. Крылова, С. И. Коржинского, К. С. Мережковского связывают первые познания в этой области. В начале XX века в Казани формируется Высшая летняя школа, в которой не на последнее место ставились вопросы фитопатологии. На протяжении многих лет активно работали такие ученые как А. А. Юницкий, А. М. Алексеев, Н. М. Ведерников, Н. П. Арискина, В. В. Гуляев и многие другие. С ХХ века начинается изучение лихенизированных грибов Татарстана исследователями К. С. Мережковским, Н. В. Малышевой, А. С. Евстигнеевой, Э. И Байбаковым, Г. П. Урбанавичюсом.
Территория Республики Татарстан находится на месте слияния двух рек – Волги и Камы. Разнообразие растительного и животного мира объясняется двумя ландшафтными зонами граничащими друг с другом. Более 17% территории покрыты лесами, в которых обитают таежные и степные виды растений и животных.
С Николая Васильевича Сорокина, а точнее с его работы «Микологические исследования» в 1872 году начинаются первые сведения о грибах Республики Татарстан. С того времени начинается описание биоразнообразия грибов Казанской губернии. В опубликованном Н. В. Сорокиным списке видов упоминается о Boletusedulis и B. luridus, которые, как он сообщает, «продаются на рынке и приносятся из ближних лесов», кроме того в список попали некоторые паразитические виды насекомых и растений. В списке сборов Зилантовского монастырь, ботанического сада, территории поселка Бутырки приводятся сведения о 157 видах грибов. Были найдены паразиты животных и растений, сапротрофы, в том числе ксило- и копротрофы, а также грибы-микоризообразователи. Н. В. Сорокин пишет «Несмотря на важность и пользу, которую имеют списки растений известной местности, на долю грибов выпала самая грустная участь...» [Бондарцева, Пармасто, 1986]. Прошло почти полтора века, а его сведения в отношении некоторых регионов по-прежнему остаются актуальными, что говорит о невысокой скорости исследований в сфере микологии.
Сборы П. Н. Крылова, собранные им в 1873 году в черте Казани, хранятся в гербарии Ботанического музея Казанского Федерального Университета. Почти в это же время, в 1882-1884 гг. С. И. Коржинский и П. Н. Крылов занимались сборами лишайников, определением которых чуть позже занимался А. А. Еленкин. Они вошли в труд из нескольких частей «Флора лишайников Средней России», которые были изданы в 1906, 1907 и 1911 гг., где автор приводит 92 вида лишайников на территории Татарстана.
В 1885 году выходит работа Сергея Ивановича Коржинского, под названием «Uredineae Казанской губернии», во введении которой пишет «По поручению Казанского Общества Естествоиспытателей летом 1884 года я экскурсировал в Казанской губернии для собирания материалов к изучению ее флоры споровых. Посетив все уезды Казанской губернии, кроме Спасского, я имел возможность собрать в числе прочих споровых также коллекцию грибов» [Бондарцев, Зингер, 1950] Работа на 25 страниц, представляет интересные сведения в исторической реконструкции и изучении изменений биоразнообразия региона.
С 1902 года Константин Сергеевич Мережковский известный русский ученый, биолог начинает работать в Казанском университете. Им была собрана коллекция лишайников, которая по сей день хранится в KAZ. Так же он опубликовал немалое количество работ, связанных с исследованиями в области лихенологии.
Среди работ 20 - 30-е годов ХХ века встречаются: «Температурный режим снежного покрова и развитие снежного шютте на сеянцах сосны», «Зараженность лиственных лесов грибными вредителями», «Важнейшие грибные вредители лесов Казанского края» и ряд других [Васильева, 1977; Гордиенко, 1979, Давыдкина, 1980], что говорит о возрастании интереса к макро- и микроскопическим грибам.
Из интереса к лесному опытному делу был порожден интерес к фитопатологии. А. А. Юницкий является основоположником фитопатологической школы Поволжья. На протяжении многих десятилетий его книги пользовались высокой ценностью и точностью информации. В дальнейшем, вплоть до 70-х гг. XX столетия в данном направлении работают Н. П. Арискина, В. В. Гуляев, Н. М Ведерников, А. Ф. Григорьев, П. Дворжецкий, В. С. Ермилова, П. Г. Трошанин и другие [Каламеэс, 1965; Гордиенко, 1979; Ивантер, Кузнецов, 2007; Малышева Е., Малышева В., 2008; Змитрович, 2008; Мюллер, Лёффлер 1995]. Публикации этого времени освещают о грибных заболеваниях, причинах возникновения и распространения сердцевидной гнили черной ольхи и осины, а так же способы противостояния ей и т.д.
В 1927 году А. М. Алексеев пишет статью «Материалы к микологической флоре Татарской Республики» [Ставишенко, Лугнина, 2015], в которой заключает, что ранние сведения работ Н. В. Сорокина и С. И, Коржинского очень скудны. Далее он пишет «Желая пополнить сведения о грибной флоре Татарской Республики, я летом 1924 и 1925 г. производил сборы грибов в ближайших окрестностях г. Казани… Обработку собранного материала я производил отчасти в Казани в Кабинете сельско-хозяйственной фитопатологии Казанского института сельского хозяйства и лесоводства, отчасти в Ленинграде в Микологической и фитопатологической лаборатории имени А. А. Ячевского…» [Ставишенко, Лугнина, 2015]. В своей работе он дает информацию о 221 виде грибов, грибоподобных протистов и некоторых слизевиках. 129 видов из данного списка являлись новыми для республики, а 4 вида вошли - как новые для России.
В 1938 – 1940 гг., в то время на территории «Раифского леса» нынешнего Волжско - Камского природного биосферного заповедника Васильева Любовь Николаевна собирает микологический материал. Список сборов ограничивается двумя классами грибов - Ascomycetes и Basidiomycetes в сумме давшие 671 вид, что в свою очередь составляет 60% % всех нелехинизированных грибов, обнаруженных за весь период изучения видового разнообразия макромицетов Республики Татарстан.
В 80-е Г. А. Юпина начинает работать на территории Волжско – Камского заповедника и прилегающих к нему территориях. В своей диссертации на тему «Экология афиллофоровых грибов лесных биогеоценозов Волжско - Камского заповедника и прилегающих антропогенных территорий» даёт информацию о видах трутовых и кортициоидных грибов, которые пополнили сведения о видовом разнообразии грибов РТ [Ставишенко, Лугнина, 2015].
Первое издание Красной Книги Татарстана выходит в 1995 году, в список которой попали 34 макро- и микроскопических вида грибов. Во втором издании указывается уже 40 редких видов грибов. Из публикаций последних 20 лет можно судить о особом внимание к охране биоразнообразия грибов.
1.5. Географическая характеристика ВКГПБЗ
С именем профессора А. Я. Гордягина связана задумка организации заповедника в 1917 г. Проект создания Татарского национального парка был внесен в Совнарком в 1924 г. Казанским институтом сельского хозяйства и лесоводства. Данный проект был утвержден Государственной комиссией, но не был реализован до 1960 г. Заповедник площадью 769 га, именовался - Волжско-Камский заповедник. Он состоит из двух участков - Раифского и Сараловского, расположенных на левобережье Волги, которые на 100 км удалены друг от друга. Территория Раифского участка площадью 3854 га располагается в 30 км западнее Казани. На 60 км южнее, в устье Камы находится Сараловская часть в 4170 га, которая помимо суши включает в себя 500 - метровую полосу прилегающей акватории Куйбышевского водохранилища, составляющая 1353 га.
Волжско-Камский государственный природный биосферный заповедник [Раифский участок]. Местоположение - Зеленодольский район РТ, в 15 км к востоку от г. Зеленодольск. Площадь - 5921 га. Склоны и днище, осложненные древними эрозионными, провальными формами рельефа, обуславливает разнообразие микроклиматических и почвенных условий, что говорит о высоком биоразнообразие. На участке выявлено сосудистых растений - 725 видов, мхов - 190, водорослей - 560, лишайников - 210, грибов - около 900, млекопитающих - 52, птиц - 170, рептилий - 6, амфибий - 10, рыб - 17, беспозвоночных – более 3000 видов. Из которых 96 видов растений, 22 вида грибов и 89 видов животных внесены в Красную книгу РТ. Стационарные исследования животного мира были начаты с 1948 г. Ведется комплексный экологический мониторинг с 1962 г. по программе «Летопись природы». Накопленная научная информация обуславливают высший природоохранный статус территории, ее познавательное значение. Имеются музей природы заповедника и дендрарий, который был заложен еще в 1921 г. Так же, для студентов функционирует стационар по проведению учебных полевых практик. На берегу озера Раифское располагается Раифский монастырь. [Государственный реестр ООПТ, 2007]
Рельеф Раифского участка равнинный, с оврагами и балками. Материковая часть Сараловского участка представляет собой выровненную поверхность, кроме северо-западного участка, в котором расположена Лысая гора. Из глины и песка сложена Материковая часть, берега которой разрушаются водохранилищем. Климат Волжско-Камского государственного природного биосферного заповедника умеренно континентальный с резкими колебаниями температуры и выпадением осадков.
К сухим рекам относят реки Раифского участка, который в летний период пересыхают. Крупные овраги и балки дают большинство притоков главной реки Сумки. По ним весной и осенью текут ливневые воды. Территория Раифского заповедника включает в себя большое количество озер и болот. Самым крупным озером считается Раифское, которое находится на месте слияния Сумки и Сер-Булака.
Почвы заповедника дерново-подзолистые. Более 90% его территории покрыты лесами. В Раифском участке на небольшой площади прослеживается географическая поясность лесной зоны Европейской части России. Если двигаться в северном направлении от южной опушки, то пройдя полосу широколиственных лесов, состоящую из липы и дуба, мы очутимся сперва в смешанных лесах, где появляется ель, а затем сосна и, в итоге, окажемся в хвойных лесах из сосны, ели и пихты, которые в свою очередь переходят в чистый сосновый лес. Возраст некоторых насаждений достигает 200-300 лет. Фауна Раифы смешана, вместе с видами широколиственных лесов обитают таежные виды животных [Васильева, 1977].
Заповедник соседствует с землями сельскохозяйственных предприятий, таких как птицефабрика, зверохозяйство, овощеводческое хозяйство. Они в свою очередь оказывают негативное воздействие на почвы, водоемы, растительный и животный мир. Например, смывается почвенный слой при весеннем таянии снега с близлежащих полей. Большая масса взвешенных частиц попадает в малые реки, которые протекающие на территории заповедника, эта масса оседает в озерах, вызывая их обмеление и сокращение площади водного зеркала. Так, Раифское озеро, глубина которого еще 20 лет назад составляла 21 м, обмелело на два метра, а площадь сократилась от 38 до 36 га. Так же, негативно влияет смыв органических веществ с полей и поступающие с территорий хозяйств неочищенные сточные воды. Чувствительно снизился сток заповедных речек Сумки и Сер-Булака, которые пересыхают на некоторых участках в летний сезон. В связи с этим начали завозить бобров, которые являются хорошими строителями гидротехнических сооружений.
Глава 2. Материалы и методы
Объектом исследования послужила биота макромицетов Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника (Рис. 3, [http://www.zapoved.net/index.php]) . Под макромицетами понимается группа грибов, имеющих макроскопические плодовые тела мясистой, хрящеватой или кожистой, пробковой консистенции.
Результатом многих адаптаций к окружающей среде являются трофические связи [Великанов, Успенская, 1980]. По мнению некоторых исследователей, основным фактором, определяющий трофическую группу является субстрат. Грибы дифференцированы на три основные группы - биотрофы, сапротрофы и симбиотрофы. Топическая характеристика неотделима от трофической, поэтому классификация А. Е. Коваленко считается наиболее оптимальной [Бондарцева, Нездойминого, 1996]. Согласно системе Коваленко 1980 г., с некоторыми изменениями [Морозова, 2001], учитываются следующие трофические группы грибов:
1. Сапротрофы. Они растут на опаде, подстилке, гумусе, древесине неразрушенной и разрушенной, корнях и погребенной в почве древесине, мхах, экскрементах и на плодовых телах грибов.
2. Симбиотрофы. Это микоризообразователи.
3. Паразиты. Бывают факультативные и облигатные на деревьях и кустарниках.
Не все грибы относятся к одной трофической группе, есть виды, которые имеют довольно широкий спектр экологических возможностей. Они могут встречаться на нескольких субстратах, поэтому такие виды грибов относят в состав различных трофических групп.
Сбор материала.
Успешная идентификация грибов главным образом обусловлена правильным сбором материала и их детальным описанием. Плодовые тела агарикоидных базидиомицетов - базидиомы, при сушке быстро теряют окраску, может измениться размер, а так же поверхность шляпки и ножки [Переведенцева, 2007].
Для точного определения агарикоидных грибов необходимо правильно их собрать, сделать детальное описание при сборе и высушить плодовые тела.
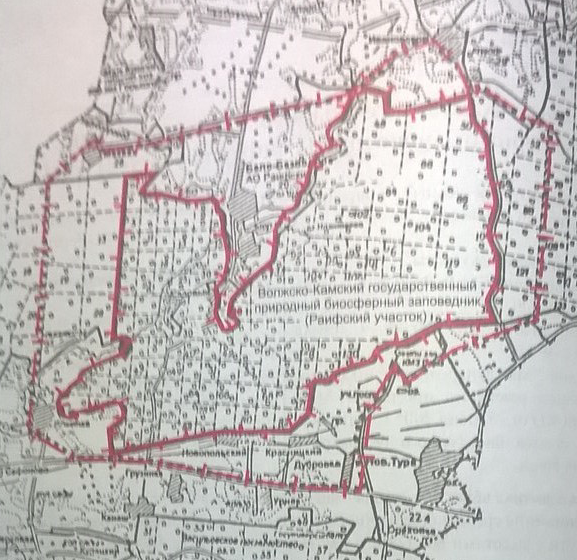
Рисунок 3. Волжско-камский государственный природный биосферный заповедник.
При сборе грибов необходимо следующее: сохранять целиком ножку, аккуратно извлекая ее из субстрата. Если грибы растут на древесине или пнях, нужно взять часть субстрата вместе с плодовыми телами. Мелкие базидиомы нужно складывать в коробочки, отдельно от крупных образцов, для того, что бы плодовые тела не повредились. Необходимо тщательнее описывать свежие плодовые тела, а так же субстрат на котором гриб произрастал. Облегчает последующее определение грибов зарисовка или фотография свежего плодового тела.
Сушка грибов является очень ответственным моментом, так как при высокой температуре грибы могут почернеть и потерять внешний облик, а при повышенной влажности сгнить. Поэту сушить грибы следует на открытом воздухе, на солнце и желательно при потоке теплого воздуха. Грибы раскладывают на металлической сетке, марле или подвешивают на ниточках над источником тепла. После сушки и описания грибы помещают в пакетик с этикеткой, на которой описаны заметки при сборе. Необходимо записывать географический пункт [область, район, поселок], название вида гриба [если удалось сразу определить], местообитание [тип растительности, дорога], субстрат и его состояние, дату, запах гриба, форму и цвет [Переведенцева, 2007].
Сбор материала осуществлялся с начала августа по конец октября 2014 года методом пробных площадей.
Снаряжение для сбора материала и данных:
1] специальный контейнер с отсеками;
2] нож с широким лезвием для выкапывания гриба или срезания его с субстрата;
3] бумага, фольга или конверт, в которую отдельно заворачивается каждый вид гриба;
4] метр для измерения длины валежа;
5] Высотомер лесотаксационный, прибор для измерения высоты стоящих деревьев.
Заложение пробных площадей происходило следующим образом.
Записывались данные географического положения пробной площади. Участок выбирался относительно однородный по своей растительности. Каждые 10-14 дней проводился повторный осмотр площадей и сбор грибов. Описанию подвергались общий характер рельефа, микрорельеф, окружение, мертвая подстилка. Описывался каждый ярус с указанием вида растения и доминирующего вида в каждом ярусе. Так же, оценен тип леса и степень сомкнутости крон. Указана формула древостоя, всходы деревьев, происхождение подроста, характер распределения подлеска, общая степень проективного покрытия всех видов в целом, а так же мохово-лишайниковый покров. Описывался древостой с указанием породы, состояния, высоты и длины окружности ствола каждого дерева. В травяно-кустарничковом ярусе было указано обилие и характер размещения каждого вида растения. В ходе работы так же проводился пересчет валежа, в описание характеристики каждой породы входило три пункта:
- стадия разложения;
- биота валежного субстрата;
- длина окружности [в двух позициях] и длина валежа.
Был собран микологический материал, при сборе которого указывался вид гриба, субстрат, ярус и характеристика субстрата, на котором он произрастал. Весь собранный материал был тщательно высушен, этикетирован и определен.
Сбор, описание и фиксация гербарного материала проводили по традиционным методикам [Бондарцев, Зингер, 1950; Бондарцев, 1953].
Сведения о встречаемости видов, хорошо распознаваемых по макропризнакам, заносились в полевой дневник на основании наблюдений в природе во время маршрутных исследований.
Ввиду большого разнообразия местообитаний, удаленности и труднодоступности ряда участков, предпочтение при проведении полевых работ было отдано методу пробных площадей. Избранные площади посещались неоднократно в течение всего вегетационного периода. Это позволило выявить не только видовой состав макромицетов, но и установить приуроченность ряда видов к определенным местообитаниям, с целью выяснения или уточнения некоторых экологических параметров, наблюдать за сезонной динамикой плодоношения.
Идентификация образцов осуществлялась на кафедре ботаники КГУ с использованием световых микроскопов БМС-1, МБИ-11. Сбор, описание и фиксация гербарного материала проводили по традиционным методикам [Бондарцев, 1953; Кутафьева, 2003]. В работе широко использовали определители, монографии по отдельным таксонам и критические статьи.
В часности, определители по всем таксонам - Nordic Macromycetes [1992]; Moser [1978];
пор. Aphyllophorales - Бондарцева, 1998;
сем. Bolbitaceae - Watling, 1982;
сем. Boletaceae, Gomphidiaceae, Paxillaceac - Watling, 1970;
сем. Cortinariaceae- Нездойминого, 1996;
сем. Hydrophpraceae - Коваленко, 1989;
сем. Pluteaceae - Orfot, 1986;
сем. Sfrophariaceae - Watling, Gredory, 1987;
сем. Tricholomataceae p.p - Watling, Turnbull, 1998;
род Cletocybe - Hannaja, 1969;
род Coprinus - Orfon et al., 1979;
род Crepidotus - serai - Jrlet, 1995;
род Entoloma - Nooradeloos, 1991’
род Lactarius - Heilmann-Ceausenet al., 1998;
род Mycena - Maas Geesteranus, 1 992b;
род Psathyrella- van Waveren, 1985;
род Russula - Romangnesi, 1967.
Таксоны в конспекте расположены по системе, принятой в 8-ом издании «Словаря грибов Айнсворта и Бисби» [Hawks-Werth et al., 1995]. Сокращения авторов даны в соответствии с рекомендациями работы «Авторы названий грибов» [Kirk, Ansell, 1992].
При проведении анализа видового состава изучаемой микобиоты был использован ряд общепринятых статистических методов в сравнительной флористике [Шмидт, 1980].
При анализе эколого-трофической структуры взята шкала трофических групп, предложенная А. Е. Коваленко [1980]. Определение принадлежности грибов к определенной трофической группе, а также установлении древесного симбиота проводилось как по личным наблюдениям, так и с использованием литературных данных [Селиванов, 1980; Шубин, 1988; Curtecuisse, Duhem, 1996].
Для выявления географической структуры использована система региональных географических элементов, с учетом некоторых экологических особенностей видов.
Глава 3. Результаты исследования
3.1. Систематическая структура.
Анализ любой конкретной флоры [микобиоты] подразумевает выявление степени богатства или бедности биоразнообразия путем изучения таксономической структуры; ее место в системе других микобиот обнаруживается после установления географической структуры; консортивные связи выявляются после анализа и трофических особенностей. В заключении анализируются ресурсная структура и созологические аспекты. В работе мы придерживались данной системы анализа.
Данный элемент анализа, выявляя ведущие таксоны, даёт возможность судить в сравнительном аспекте о степени биологического разнообразия.
Аннотированный перечень макромицетов ВКГПБЗ
* - Новые виды для ВКПБЗ
** - Новые виды для РТ
Отдел ASCOMYCOTA
Класс LEOTIOMYCETES
Порядок HELOTIALES
Семейство Dermateaceae
**Catinella olivacea (Batsch) Boud. На валежном стволе сосны в сосняке с елью, раифский участок ВКГБЗ, 21.09.2014.
Семейство Helotiaceae
**Neobulgaria pura (Pers.) Petr. На валеже березы в ПП3, кв. 48. 08.09.2014.
Семейство Pezizaceae
**Pachyella subisabellina (Le Gal) Trimbach. На валеже березы в широколиственном лесу, кв.88. 18.08.2014.
Отдел BASIDIOMYCOTA
Класс AGARICOMYCETES
Порядок AGARICALES
Семейство Agaricaceae
Melanophyllum haematospermum (Bull.) Kreisel . На гнилой древесине в липняке с дубом и вязом, ПП1, кв.86, 08.09.2014;
Семейство Amanitaceae
Amanita muscaria (L.) Lam. На почве в ПП3, кв. 86, 08.09.2014.
Семейство Entolomataceae
Entoloma nitidum Quél. Во мху на почве в ПП3, кв. 86, 08.09.2014.
Семейство Hydnangiaceae
Laccaria laccata (Scop.) Cooke. На почве в липняке с дубом и вязом, ПП1, кв.86, 08.09.2014; На подстилке в ПП2, кв. 86, 08.09.2014.
Семейство Mycenaceae
Mycena acicula (Schaeff.) P. Kumm. На мелкомерных древесных остатках, ПП1, кв. 86. 08.09.2014;
Mycena epipterygia (Scop.) Gray. на подстилке в ПП3, кв. 48. 08.09.2014.
*Mycena flavoalba (Fr.) Quél. На подстилке в ПП3, кв. 48. 08.09.2014.
*Mycena haematopus (Pers.) P. Kumm. На валеже лиственной породы, ПП1, кв.86, 08.09.2014;
Mycena inclinata (Fr.) Quél. На валеже дуба в широколиственном лесу, маршрут, кв.89. 18.08.2014.
*Mycena pelianthina (Fr.) Quél. На почве в липняке с дубом и вязом, ПП1, кв. 86. 08.09.2014; На почве в ПП2, кв. 86, 08.09.2014.
Mycena pura (Pers.) P. Kumm. На подстилке в ПП2, кв. 86, 08.09.2014.
Mycena rosea (Schumach.) Gramberg. На почве в липняке с дубом и вязом, ПП1, кв. 86. 08.09.2014;
*Mycena sanguinolenta (Alb. & Schwein.) P. Kumm. На подстилке в ПП2, кв. 86, 08.09.2014.
Panellus stipticus (Bull.) P. Karst. На валеже березы в в липняке с дубом и вязом, ПП1, кв. 86. 08.09.2014;
Семейство Pluteaceae
Pluteus cervinus (Schaeff.) P. Kumm. На валеже лиственной породы в широколиственном лесу, маршрут, кв.89. 18.08.2014.
**Pluteus hongoi Singer. На валежной древесине в широколиственном лесу, маршрут, кв.89. 18.08.2014.
Pluteus leoninus (Schaeff.) P. Kumm. На валеже лиственного дерева в широколиственном лесу, маршрут, кв.87. 18.08.2014.
Pluteus romellii (Britzelm.) Lapl. На подстилке в ПП2, кв. 86, 08.09.2014.
Pluteus semibulbosus (Lasch) Quél. На мелкомерном валеже лиственной породы в широколиственном лесу, маршрут, кв.89. 18.08.2014.
Семейство Psathyrellaceae
Coprinellus micaceus (Bull.) Vilgalys. На подстилке в ПП2, кв. 86, 08.09.2014.
Psathyrella candolleana (Fr.) Maire. На почве в широколиственном лесу, маршрут, кв.89, 18.08.2014.
Порядок BOLETALES
Семейство Boletaceae
Leccinum scabrum(Bull.) Gray. На почве в ПП2, кв. 86, 08.09.2014.
Xerocomus ferrugineus (Schaeff.) Alessio. На почве в ПП3, кв. 86, 08.09.2014.
Семейство Hygrophoropsidaceae
Hygrophoropsis aurantiaca (Wulfen) Maire. На почве в ПП3, кв. 86, 08.09.2014.
Leucogyrophana mollusca (Fr.) Pouzar. на валежной ветви сосны в ПП3 (две встречи), кв. 86, 08.09.2014.
Семейство Paxillaceae
Paxillus involutus (Batsch) Fr. На почве в ПП3, кв. 86, 08.09.2014.
Порядок CANTHARELLALES
Семейство Clavulinaceae
Clavulina coralloides (L.) J. Schröt. На почве в липняке с дубом и вязом, ПП1, кв. 86. 08.09.2014;
Порядок HYMENOCHAETALES
Семейство Schizoporaceae
**Xylodon asperus(Fr.) Hjortstam & Ryvarden. На валеже сосны в ПП3, кв. 48. 08.09.2014.
Порядок PHALLALES
Семейство Phallaceae
Phallus impudicusL. var. impudicus. На почве в ПП2, кв.86. 14.08.2014.
Порядок POLYPORALES
Семейство Fomitopsidaceae
Antrodia xantha (Fr.) Ryvarden. На валеже сосны в ПП3 (две встречи), кв. 48. 08.09.2014.
Antrodia sinuosa (Fr.) P. Karst. На валеже сосны в ПП3 (две встречи), кв. 48. 08.09.2014.
Fomitopsis pinicola (Sw.) P. Karst. На сухостое березы в ПП2, кв.86. 14.08.2014.
Postia fragilis (Fr.) Jülich. на валежной ветви сосны в ПП3, кв. 48. 08.09.2014.
Postia ptychogaster (F. Ludw.) Vesterh. На валеже сосны в ПП3, кв. 48. 08.09.2014.
Семейство Polyporaceae
Hapalopilus croceus (Pers.) Donk. На валеже дуба в широколиственном лесу, маршрут, кв.87. 18.08.2014.
Порядок RUSSULALES
Семейство Auriscalpiaceae
Lentinellus vulpinus (Sowerby) Kühner & Maire. На валежной древесине в широколиственном лесу, маршрут кв.88. 18.08.2014.
Семейство Stereaceae
Stereum subtomentosum Pouzar. На валеже березы в ПП3, кв. 48. 08.09.2014.
Xylobolus frustulatus (Pers.) Boidin. На валеже дуба в широколиственном лесу, маршрут, кв.87. 18.08.2014.
Порядок THELEPHORALES
Семейство Thelephoraceae
Thelephora palmata (Scop.) Fr. На почве в ПП3, кв. 48. 08.09.2014.
Tomentella stuposa (Link) Stalpers. На валеже сосны в ПП3, кв. 48. 08.09.2014.
Класс PUCCINIOMYCETES
Порядок PLATYGLOEALES
Семейство Platygloeaceae
**Eocronartium muscicola (Pers.) Fitzp. Паразитирует на мхе, растущем на валежном стволе, кв.89, 18.08.2014.
Класс TREMELLOMYCETES
Порядок TREMELLALES
Семейство Exidiaceae
*Exidia saccharina (Alb. & Schwein.) Fr. На валеже сосны в ПП3, кв. 48. 08.09.2014.
Аннотированный список макромицетов экосистем заповедника, являющийся результатом проведенных исследований, насчитывает 45 видов, относящихся к 31 родам, 23 семействам. Из этого количества 11 видов впервые отмечены для исследуемой территории: Catinella olivacea (Batsch) Boud; Neobulgaria pura (Pers.) Petr; Pachyella subisabellina (Le Gal) Trimbach; Pluteus hongoi Singer; Xylodon asperus (Fr.) Hjortstam & Ryvarden; Eocronartium muscicola (Pers.) Fitzp; Mycena sanguinolenta (Alb. & Schwein.) P. Kumm; Mycena pelianthina (Fr.) Quél; Mycena flavoalba (Fr.) Quél; Mycena haematopus (Pers.) P. Kumm; Exidia saccharina (Alb. & Schwein.) Fr. Из них 6 видов являются новыми для Республики Татарстан: Catinella olivacea (Batsch) Boud; Neobulgaria pura (Pers.) Petr; Pachyella subisabellina (Le Gal) Trimbach; Pluteus hongoi Singer; Xylodon asperus (Fr.) Hjortstam & Ryvarden; Eocronartium muscicola (Pers.) Fitzp.
Выявленное число грибов не является исчерпывающим, так как ряд видов, встречающихся на сопредельных территориях и не указанных в данной работе, вероятнее всего встретятся в заповеднике в будущих исследованиях.
Таблица 1. Основные пропорции макромицетов Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника
|
№ |
Таксоны |
Количество |
родовой коэффициент |
|||||
|
Семейств |
Родов |
Видов |
||||||
|
число |
% |
число |
% |
число |
% |
|||
|
1 |
Ascomycètes |
3 |
20 |
3 |
24 |
3 |
5,1 |
1,5 |
|
2 |
Basidiomycetes |
20 |
80 |
28 |
76 |
42 |
94,9 |
2,9 |
|
Всего |
23 |
100 |
31 |
100 |
45 |
100 |
2,2 |
|
Родовой коэффициент, определяющийся отношением числа видов к числу родов, составляет 2,2. Абсолютно в количественном отношении преобладают представители класса Basidiomycetes, содержащие 42 вида из 28 родов, 20 семейств, с родовым коэффициентом 2,9. Всего 3 видами представлен класс Ascomycetes из 3 родов, 3 семейств [табл.1].
Среднее содержание видов в семействах макромицетов отдела Basidiomycetes составляет 7,9; Ascomycetes - 2,2, соответственно, в общем - 5,05. Лишь 7 семейств достигают такой видовой насыщенности. Два ведущих семейств содержат в общей сложности 15 видов или 34,2% от общего количества [табл.2, рис.4]. Как видно из данной таблицы ведущими семействами являются: Tricholomataceae, Cortinariaceae, Russulaceae. Этот факт подчеркивает бореальный характер исследуемой микобиоты. Обилие видов семейства Agaricaceae составляющий 8,9% от всей микобиоты бывшей территории СССР [Вассер, 1984], так же свидетельствует о бореальном характере. Доминирующее положение этих трех семейств характерно также для близ лежащих территорий. Они занимают господствующие положение в сложении микобиоты бореальных зон.
Таблица 2. Ранжирование ведущих семейств
|
№ |
Семейство |
Количество видов |
|
|
число |
% |
||
|
1 |
Mycenaceae |
10 |
22,8 |
|
2 |
Fomitopsidaceae |
5 |
11,4 |
|
3 |
Psathyrellaceae |
2 |
4,56 |
|
4 |
Boletaceae |
2 |
4,56 |
|
5 |
Hygrophoropsidaceae |
2 |
4,56 |
|
6 |
Stereaceae |
2 |
4,56 |
|
Итого |
23 |
51,1 |
|
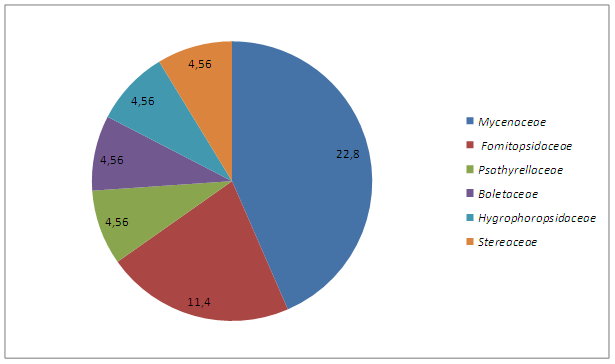
Рисунок 4. Спектр ведущих семейств макромицетов
Для выявления особенностей микобиоты необходим анализ родового спектра. Как видно из табл.1, всего в микобиоте макромицетов зарегистрирован 31 род. Это количество родов составляют в общей сложности 45 видов [100%].
Спектр ведущих 8 родов в убывающем порядке представлен следующим реестром: Мусепа –> Pluteus —> Antrodia —> Postia.
Количественный состав каждого рода представлен в таблице 3 и рисунке 5.
Таблица 3. Ранжирование ведущих родов
|
Рода |
Количество видов |
||
|
N |
число |
% |
|
|
1 |
Мусеnа |
9 |
19,63 |
|
2 |
Pluteus |
5 |
12,10 |
|
3 |
Antrodia |
2 |
10.90 |
|
4 |
Postia |
2 |
4.36 |
|
Итого: |
18 |
40 |
|
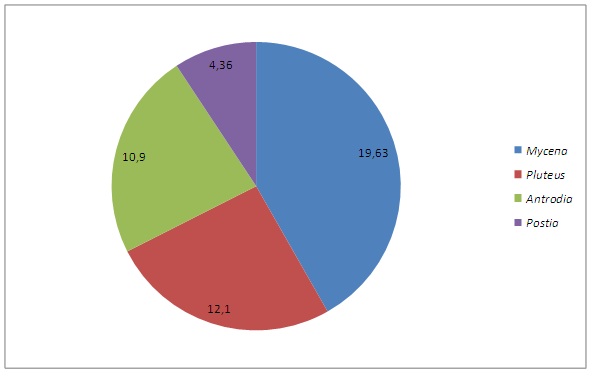
Рисунок 5. Спектр ведущих родов
Большиство крупных родов относятся к крупным семействам: Mycenaceae -> Mycena; Pluteaceae -> Pluteus; Fomitopsidaceae —> Antrodia, Postia [табл.4]. Как видно из данной таблице в родово - видовом спектре ведущее место среди порядков занимают Agaricales, Boletales и Polyporales содержащие 30 видов [67%]. Средних родов, насчитывается 23 вида в микобиоте содержащие 15 [23%] видов.
Виды, встречающиеся более 1 раза представлены в таблице 4.
Таблица 4. Повторяющиеся виды.
|
№ |
Виды |
Число грибов |
№ |
|
1 |
Laccaria laccata (Scop.) Cooke |
2 |
4,08 |
|
2 |
Mycena pelianthina (Fr.) Quél |
2 |
4,08 |
|
3 |
Leucogyrophana mollusca (Fr.) Pouzar |
2 |
4,08 |
|
4 |
Antrodia xantha (Fr.) Ryvarden |
2 |
4,08 |
|
5 |
Antrodia sinuosa (Fr.) P. Karst |
2 |
4,08 |
|
Итого |
10 |
20,40 |
|
Всего в ходе данной работы было собрано 49 грибов, при определении которых были выявлены повторяющиеся виды, а именно: Laccaria laccata; Mycena pelianthina; Leucogyrophana mollusca; Antrodia xantha; Antrodia sinuosa. В сумме 5 видов составили 10 грибов [20,4%] от всего количества собранных образцов.
Сводная таксономическая структура микобиоты макромицетов экосистем Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника приведена в таблице 5.
Таблица 5. Таксономическая структура макромицетов Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника
|
Отдел |
Класс |
Порядок |
Семейство |
Род |
Число видов |
|
Ascomycota |
Leotiomycetes |
Helotiales |
Dermateaceae |
Catinella |
1 |
|
Helotiaceae |
Neobulgaria |
1 |
|||
|
Pezizaceae |
Pachyella |
1 |
|||
|
Basidiomycota |
Agaricomycetes |
Agaricales |
Agaricaceae |
Melanophyllum |
1 |
|
Amanitaceae |
Amanita |
1 |
|||
|
Entolomataceae |
Entoloma |
1 |
|||
|
Hydnangiaceae |
Laccaria |
1 |
|||
|
Mycenaceae |
Mycena |
9 |
|||
|
Panellus |
1 |
||||
|
Pluteaceae |
Pluteus |
5 |
|||
|
Psathyrellaceae |
Coprinellus |
1 |
|||
|
Psathyrella |
1 |
||||
|
Boletales |
Boletaceae |
Leccinum |
1 |
||
|
Xerocomus |
1 |
||||
|
Hygrophoropsidaceae |
Hygrophoropsis |
1 |
|||
|
Leucogyrophana |
1 |
||||
|
Paxillaceae |
Paxillus |
1 |
|||
|
Cantharellales |
Clavulinaceae |
Clavulina |
1 |
||
|
Hymenochaetales |
Schizoporaceae |
Xylodon |
1 |
||
|
Phallales |
Phallaceae |
Phallus |
1 |
||
|
Polyporales |
Fomitopsidaceae |
Antrodia |
2 |
||
|
Продолжение таблицы 5 |
|||||
|
Отдел |
Класс |
Порядок |
Семейство |
Род |
Число видов |
|
Basidiomycota |
Agaricomycetes |
Polyporales |
Fomitopsidaceae |
Fomitopsis |
1 |
|
Postia |
2 |
||||
|
Polyporaceae |
Hapalopilus |
1 |
|||
|
Russulales |
Auriscalpiaceae |
Lentinellus |
1 |
||
|
Basidiomycota |
Stereaceae |
Stereum |
1 |
||
|
Xylobolus |
1 |
||||
|
Thelephorales |
Thelephorales |
Thelephora |
1 |
||
|
Tomentella |
1 |
||||
|
Pucciniomycetes |
Platygloeales |
Platygloeaceae |
Eocronartium |
1 |
|
|
Tremellomycetes |
Tremellales |
Exidiaceae |
Exidia |
1 |
|
3.2 Трофическая структура
Субстрат - один из важных экологических факторов в жизни макромицетов так как, являясь гетеротрофными организмами, они питаются, получая необходимые питательные вещества из него. Этот факт давно замечен микологами, а потому является одним из элементов анализа микобиоты. Были предложены различные классификации субстрата макромицетов. Наиболее удачное и принятое название «трофические группы». По типу питания все выявленные макромицеты района исследований подразделяются на симбиотрофы [Мг-микоризообразователи] и сапротрофы. Сапротрофные грибы по приуроченности к определенным разновидностям субстрата согласно А. Е. Коваленко [1980], отнесены к следующим группам: на опаде [Fd], на подстилке [St], на гумусе [Нu], на разрушенной древесине [Lep], на неразрушенной древесине [Lei], на корнях деревьев и погребенной в почву древесине [Lh], копротрофы [Е], карботрофы [С], на мхах [Ms]. В итоге по типу питающего субстрата выявленные макромицеты отнесены к десяти трофическим группам [табл.6 и рис.6], характеристики которых даем ниже.
Сапротрофы на опаде [Fd] - объединяют группу макромицетов заселяющие растительный опад полностью сохранивший структуру [прошлогодние листья, хвоинки, мелкие веточки], отдельные компоненты лежат свободно и не связанны гифами мицелия. Эта группа включает 2 вида относящихся к 1 роду и 8 семействам.
Таблица 6. Трофические группы в микобиоте экосистем Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника
|
Трофические группы |
Количество видов |
|
|
число |
% |
|
|
Симбиотрофы Микоризообразователи [Мr] |
37 |
39,79 |
|
Сапроарофы на: |
39 |
42,21 |
|
опаде [Fd] |
2 |
2,1 |
|
подстилке [St] |
2 |
2,1 |
|
гумусе [Нu] |
2 |
2,1 |
|
разрушенной древесине [Lep] |
1 |
1,05 |
|
неразрушенной древесине [Lei] |
1 |
1,05 |
|
погребенной древесине [Lh] |
1 |
1,05 |
|
копротрофы [Е] |
1 |
1,05 |
|
карботрофы [С] |
2 |
2,1 |
|
мхах [Ms] |
2 |
2,1 |
Сапротрофы на подстилке [St] - это макромицеты заселяющие слой частично разрушенного растительного опада, плотно переплетенного гифами мицелия. Представители данной эколого-трофической группы выполняют функцию минерализации и гумификации органических веществ, образующихся в результате разложения растительного опада. Группа включает 2 вида относящиеся к 1 роду и 1 семейству Tricholomataceae.
Гумусовые сапротрофы [Нu] - группа грибов, мицелий которых расположен в гумусовом горизонте почвы и осуществляет его разложение. Гумусовый слой почвы благодаря специфичному составу органических веществ, однородностью механического состава, относительной стабильности экологических и биохимических показателей древесине разной степени разложения представляет собой специфическую среду обитания грибов. Она включает 2 вида относящихся к 1 роду и 1 семейству Agaricaceae.
Сапротрофы на древесине [ксилотрофы] - макромицеты обитающие на древесине разной степени разложения, использующие ее биополимеры [целлюлозу и лигнин] в качестве питания. По мере разрушения древесины изменяются ее физико-химические свойства [плотность, влажность, термический режим, химизм]. Чем и обуславливается смена грибов-диструктиров. При этом одни виды заселяют древесину вскоре после ее отмирания [группа Lei], другие на более поздних стадиях [группа Lep]. Часть же грибов поселяются на живых или ослабленных деревьях [Le]. Другая часть макромицетов обитают на древесине любой стадии разрушения, включая стадию древесной «трухи» и способны осуществлять полную деструкцию древесины.
Разделение древесины на разрушенную и не разрушенную, достаточно условно. В данной работе разграничения приводим в соответствии с представлениями А. Е. Коваленко [1980]:
· не разрушенная древесина- древесина от момента отмирания до момента утраты коры и размягчения верхнего слоя.
· разрушенная древесина- древесина от момента частичной утраты коры и размягчения, до полного размягчения и распада на отдельные куски.
К разрушенной древесине относятся и небольшие гнилушки, сохраняющие структуру древесины. Наиболее благоприятными для развития макромицетов являются крупные валеженные бревна и пни.
Копротрофы [E] – макромицеты, трофически связанные с пометом, главным образом домашних травоядных животных. В основном встречаются вдоль лесных дорог, на полянах, опушках.
Сапротрофы на мхах [Ms] – виды, связанные с зелеными мхами. Группа содержит 2 видов [1,05%] из следующих семейств: Tricholomataceae, Cortinariaceae. Это: Laccaria bicolor, L laccata, Galera hypnorum.
Симбиотрофы [микоризообразователи Мг] - грибы, образующие микоризу с древесными и кустарниковыми растениями. Проведенные исследования показывают, что наибольшее число видов макромицетов на изучаемой территории связано трофически с основной лесообразующей породой района исследований Fagus orientalis. Бук является самой распространенной древесной породой и эдификатором сообществ, а так же встречается в качестве примеси во многих ассоциациях, образованных другими породами деревьев. Так же трофически связаны с представителями рода Quercus, Carpinus, Betula. Группа симбиотрофов содержит 37 вида [39,79%].
Ведущими семействами в группе являются: Russulaceae [13 видов, 16,45%], Tricholomataceae [11 видов, 13,92%], Boletaceae [9 видов, 11,39%], Cortinariaceae [8 видов, 10,12%].
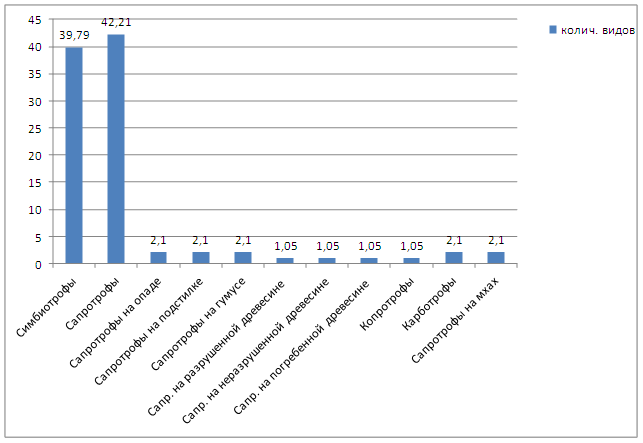
Рисунок 6. Соотношение трофических групп в биоте макромицетов Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника.
Политрофы - Pt, сапротрофы на опаде - Fd, сапротрофы на подстилке - St, гумусовые сапрофиты Hu, сапротрофы на разрушенной древесине - Lep, сапротрофы на неразрушенной древесине - Lei, сапротрофы на погребенной древесине - Lh, капротрофы - Е, карботрофы- C, сапротрофы на мхах - М, симбиотрофы [мнкорнзообразователи].
Политрофы грибы способные разлагать два и более субстрата. Их трофические группы представляют собой комбинации моносапротрофных и симбиотрофных групп. Большинство видов способны разлагать 2-3-4 субстрата.
Таким образом, трофическая структура биоты макромицетов экосистем Раифского участка Волжско-Камского Государственного Природного Биосферного Заповедника представлена 10 трофическими группами. Основными функциями микобиоты является разложение органических веществ растительного происхождения. Из них преобладают гумусовые сапрофиты.
Они включают 45 видов из 31 рода и 23 семейств. Наибольшую активность проявляют семейства Agaricaceae и Сорппасеае. Значительное количество видов также на опаде [26 видов] и подстилке [24 вида].
Политрофность [индифферентность] показывают более 19 видов, встречаясь на 2-3-4 субстратах. Так, установлено, что на двух субстратах встречается 52,53% видов; на трех -17,25%; на четырех- 4,56%.
Выводы
В результате проведенных исследований биоты макромицетов лесных экосистем Раифского участка Волжско-Камского государственного природного биосферного заповедника выявлено 45 видов, относящихся к 31 роду, 23 семействам. Среди них 11 видов впервые зарегистрированы для исследуемой территории, из них 6 видов являются новыми для Республики Татарстан.
Спектр ведущих по числу видов семейств возглавляют Mycenaceae, Fomitopsidaceae, Psathyrellaceae, Boletaceae, Hygrophoropsidaceae, Stereaceae, которые составляют 51,1% от всего количества видов, что свидетельствует о бореальном характере биоты макромицетов исследуемой территории.
В родово-видовом спектре наиболее многочисленны типично «лесные рода»: Мусеnа, Pluteus, Antrodia, Postia, содержащие в общей сложности – 40%.
Выявленные при определении повторяющимися видами оказались: Laccaria laccata; Mycena pelianthina; Leucogyrophana mollusca; Antrodia xantha; Antrodia sinuosa.
Трофическая структура микобиоты представлена 10 трофическими группами. При этом грибы сапрофитного типа питания [56,21% из 8 трофических групп] преобладают над симбиотрофами. К типу субстрата 74,34% видов изучаемой микобиоты политрофны. На двух субстратах [опад, подстилка,] отмечены 52,53% видов; на трех [опад, подстилка, гумус,] - 17,25%; на четырех [опад, подстилка, гумус, древесина] - 4,56%.Наибольшее число видов макромицетов в районе исследования трофически связаны с основными лесообразующими породами – сосна, ель, береза, дуб и липа.
Список используемых источников
1. Алексеев, А. М. Материалы к микологической флоре Татарской республики [Текст] / А. М. Алексеев // Изв. Казан, ин-та сел. хоз-ва и лесоводства. – 1927. - №1. – С. 60-97.
2. Алимова, Ф. К. Биологическое разнообразие видов рода Trichoderma [Fungi, Ascomycetes, Hypocreales] и их роль в функционировании микробиоты и защите растений в агроценозах различных почвенно-климатических зон на территории Республики Татарстан [Текст]: дис. д-ра. биол. наук: 03.00.07, 06.01.11: утв. 13.10.06 / Алимова Фарида Кашифовна. – Казань, 2006. – 435 с.
3. Атлас Республики Татарстан [Текст] / ред. Б. Г. Петрова. – СПб.: Иван Федоров, 2005. – 215 с.
4. Балашова, Н. Б. Летняя практика по альгологии и микологии в Санкт-Петербургском университете [Текст] / Н. Б. Балашова, А. В. Тобис, Д. Е. Гимельбрант. – СПб.: Учебное пособие, 2005. - 236 с. – ISBN 5-28803-770-1.
5. Билай, В. Н. Основы общей микологии [Текст] / В. Н. Билай:. Киев, Головное издательство издательского объединения «Выща школа», 1989. 360 с.
6. Бондарцева, М. А. Определитель грибов России. Порядок Афиллофоровые [Текст] / М. А. Бондарцева. Под ред. А. Е. Коваленко. – СПб.: Наука, 1998. – 391 с.
7. Бондарцева, М. А., Определитель грибов СССР. Порядок Афиллофоровые [Текст] / М. А. Бондарцева, Э. Х. Пармасто. – Вып 1.– Л.: Наука, 1986. – 191 с.
8. Бурова, Л. Г. Загадочный мир грибов [Текст] / Л. Г. Бурова – М.: Наука, 1991. – 96 с.
9. Бурова, Л. Г. Экология грибов макромицетов [Текст] / Л. Г. Бурова – М.: Наука, 1986. – 222 с.
10. Васильева, Л. Н. Грибы макромицеты Раифского участка Волжско-Камского заповедника [Текст] / Л. Н. Васильева // Труды Волжско-Камского государственного заповедника: сб. науч. тр. – Казань: Татарское книжное издательство, 1977. – Вып. 3. – С. 3–60.
11. Ведерников, Н. М. Температурный режим снежного покрова и развитие снежного шютте на сеянцах сосны [Текст] / Н. М. Ведерников // Сб. трудов по лесному хозяйству: сб. науч. тр. – Казань, 1967. – Вып. 17 – С. 241–252.
12. Гордиенко, П. В. Влияние субстрата на расселение дереворазрушающих грибов в лесах Сихоте-Алиня // Микология и фитопатология. 1979. – Т. 13. – Вып. 5. – С. 421-428.
13. Государственный реестр ООПТ в РТ. Издание второе [Текст]. – Казань: Издательство «Идел-Пресс», 2007. – 185-186 с.
14. Григорьев, А. Ф. Зараженность лиственных лесов грибными вредителями (Черемшанское лесничество ТАССР) [Текст] / А. Ф. Григорьев // Труды Общества изучения Татарстана: сб. науч. тр. – Казань: 1930. – Т.3. – С. 3-50.
15. Давыдкина, Т. А. Стереумовые грибы Советского Союза [Текст] / Т. А. Давыдкина. –Л.: Наука, 1980. – 142 с.
16. Демина, Г. В. Николай Васильевич Сорокин, 1846-1909 [Текст] / Г.В. Демина. – Казань: Изд-во Казанск. ун-та, 2006. – 16 с.
17. Дьяков, Ю. Т. Занимательная микология [Текст] / Ю. Т. Дьяков; Шедевры научно - популярной литературы. – Вып. 65. - М.: Книжный дом «ЛИБРОКОМ», 2013. – С. 8-21. - ISBN 5-39703-209-4.
18. Евстигнеева, А. С. Таксономический анализ лихенофлоры Республики Татарстан [Текст] / А. С. Евстигнеева, В.Ю. Евстигнеев // Учен. зап. Казан. ун-та. Сер. Естеств. науки, 2009. – Т. 151, кн. 2. – С. 212–223.
19. Змитрович, И. В. Определитель грибов России. Порядок Афиллофоровые [Текст] / И. В. Змитрович. Вып. 3 / Под. ред. М. А. Бондарцевой. – СПб.: Товарищество научных изданий КМК, 2008. – 278 с.
20. Змитрович, И. В. Макромицеты [Heterobasidiomycetes; Aphyllophorales - I]: Аннотированные списки видов [текст] / И. В. Змитрович // Грибы Нижнесвирского заповедника. -Вып. 3. 1999. – 66 с.
21. Каламеэс, К. А. Основные проблемы и методика микоценологических исследований [Текст] / К. А. Каламеэс // Проблемы изучения грибов и лишайников. Тарту, 1965. – С. 14 – 21.
22. Коржинский, С. И. Uredineae Казанской губернии [Текст] / С. И. Коржинский. – Казань: Типография Императорскаго Университета, 1885. – 25 с.
23. Красная книга Республики Карелия [Текст] / Ред. Э. В. Ивантер, О. Л. Кузнецов. - Петрозаводск: Карелия, 2007. - 368 с.
24. Красная книга Республики Татарстан: животные, растения, грибы [Текст] / Гл. ред. А. И. Щеповских. – Казань: Природа, 1995. – С. 424–446.
25. Красная книга Республики Татарстан: животные, растения, грибы [Текст] / Гл. ред. А. И. Щеповских. – Казань: Идел-Пресс, 2006. – С. 747–790.
26. Малышева, В. Ф. Высшие базидиомицеты лесных и луговых экосистем Жигулей [Текст] / В. Ф. Малышева, Е. К. Малышева – М.: – СПб.: Товарищество научных изданий КМК, 2008. – 242 с.
27. Мирчинк, Т. Г. Почвенная микология [Текст] / Т. Г. Мирчинк: - М.: издательство МГУ, 1988. – 220 с. – ISBN 5-21100-157-5.
28. Морозова, О. В. Агарикоидные базидиомицеты подзоны южной тайги Ленинградской области [Текст] : Дис. … канд. биол. наук / О. В. Морозова. – СПб.: БИН РАН, 2001. – 250 с.
29. Мюллер Э. Микология [Текст]: [пер. с нем.] / Эмил Мюллер, Вольфган Лёффлер — М.: Мир, 1995. — 343 с. – Перевод изд: Mykologie. /Emil Muller, Wolfgang Loeffler. - New York: Georg Thieme Verlag Stuttgart, 1992. – 343 c. - ISBN 5-03002-999-0.
30. Нездойминого, Э. Л. Определитель грибов России. Вып. 1. Порядок Агариковые. Семейство Паутинниковые [Текст] / Э. Л. Нездойминого / Отв. ред. М. А. Бондарцева. – СПб.: Наука, 1996. – 408 с.
31. Переведенцева, Л. Г. Некоторые аспекты мониторинга агарикоидных базидиомицетов в лесных ценозах Центрального Прикамья [Текст] / Л. Г. Переведенцева // Грибные сообщества лесных экосистем. Москва-Петрозаводск: Карельский научный центр РАН, 2000. – 321 с.
32. Переведенцева, Л. Г. Биоразнообразие и экология низших растений. Методика сбора, описание и определения агарикодных базидиомицетов: метод. указания для летней полевой практики [Текст] / Л. Г. Переведенцева. –Пермь: Изд-во Пермск. ун-та, 2007.-28 с.
33. Потапов, К. О. Трутовые грибы национального парка «Нижняя Кама» [Текст] / К. О. Потапов // Ботанические заметки. – 2013. – №4. – С. 30-36.
34. Потапов, К. О. Новые виды афиллофороидных и гастероидных базидиомицетов на территории Республики Татарстан [Текст] / К. О. Потапов // Вопросы общей ботаники – традиции и перспективы: сб. тр. II Международной Интернет-конф. – Казань: издательство «Казанский университет», 2011. – С. 110-114.
35. Ребриев, Ю. А. Гастероидные базидиомицеты Нижнего Дона (в пределах Ростовской области) [Текст] : Автореф. дис. … канд. биол. наук / Ю. А. Ребриев; – Ростов-на-Дону, 2002. – 18 с.
36. Сорокин, Н. В. Микологические исследования [Текст] / Н. В. Сорокин. – Казань: лито-тип. К. А. Тилли, 1874. – 50 с.
37. Сосин, П. Е. Определитель гастеромицетов СССР [Текст] / П. Е. Сосин. — Л.: «Наука», 1973. – 164 с.
38. Спирин, В. А. Афиллофоровые грибы Нижегородской области: видовой состав и особенности экологии [Текст] : автореф. дис. … канд. биол. наук / В. А. Спирин; – СПб., 2003. – 27 с.
39. Ставишенко, И. В. Микология и фитопатология [Текст] / И. В. Ставишенко, А. Е. Лугинина. – Т. 49. – Вып. 1. – 2015.
40. Черепанова, Н. П. Систематика грибов [Текст] / Н. П. Черепанова / Учеб. пособие. – 2-е изд. – СПб.: Изд-во С.-Петерб. ун-та, 2005. – 344 с.
41. Шварцман, С. Р. Гастеромицеты – Gasteromycetes. Флора споровых растений Казахстана [Текст] / С. Р. Шварцман, Н. М. Филимонова. Отв. ред. С. Р. Шварцман. – Алма-Ата: Наука, 1970. – 319 с.
42. Юницкий, А. А. Важнейшие грибные вредители лесов Казанского края [Текст] / А. А. Юницкий // Дневник Всесоюзного съезда ботаников в Ленинграде в янв. 1928 года: сб. науч. тр. – Л.: Изд. Гос. рус. ботан. о-ва, 1928. – С. 191-192.
43. Юпина, Г. А. Дереворазрушающие грибы лесных биогеоценозов Волжско-Камского заповедника [Текст] / Г. А. Юпина // Микология и фитопатология – 1987. – Т. 21, – Вып. 1. – С. 82-84.
44. Mycobank database/ Fungal Databases, Nomenclature & Species Banks [Электронный ресурс]. – Режим доступа: http://www.mycobank.org – Дата доступа: 12.02.2015.
 (zip - application/zip)
(zip - application/zip)