Міністерство освіти і науки України
Львівський національний університет імені Івана Франка
Геологічний факультет
Кафедра історичної геології та палеонтології
КУРСОВА РОБОТА
На тему:
Біостратиграфічна характеристика примежових відкладів силуру і девону Волино-Поділля за фауною трилобітів
студентки 5 курсу групи ГЛГ-5м
напрямку підготовки 6.040103
спеціальності «Геологія»
Яцкевич Ілони Михайлівни
Керівник__________________
доц., канд. геол.-мін. наук
Іваніна Антоніна Валентинівна
Національна шкала________________________
Кількість балів:__________ Оцінка: ECTS_____
Члени комісії ___________________ ____________________
___________________ ____________________
___________________ ____________________
Львів-2016
Зміст:
Вступ……………………………………………………..........................3
1. Історія визначення межі між силуром і девоном……………………4
2. Палеонтологічна характеристика відкладів верхнього силуру головних стратотипових розрізів світу і Волино-Поділля………………..…...11
3. Палеонтологічна характеристика відкладів нижнього девону головних стратотипових розрізів світу і Волино-Поділля…………………...18
4. Морфологічний опис деяких видів трилобітів примежових відкладах силуру і девону Волино-Поділля…………………………………….23
Висновки…………………………………………………………..….…28
Список використаної літератури……………………………………..29
Додаток 1…………………………………………………………….….31
Додаток 2………………………………………………………………..32
Додаток 3…………………………………………………………….….33
Додаток 4…………………………………………………….…………34
Вступ
Актуальність. Трилобіти нижньодевонських і верхньосилурійських відкладів Поділля погано вивчені. Уточнення стратиграфії палеозою Східноєвропейської платформи, кореляція нижньодевонських і верхньосилурійських відкладів з одновіковими відслоненнями інших районів колишнього СРСР і закордонних країн вимагають подальшого вивчення фауни, в тому числі трилобітів Поділля. Займаючись трилобітами північно-західної частини Російської платформи, у 1964, 1966 і 1967 рр. під час літолого-палеонтологічного вивчення нижнього девону і силуру Поділля, проведеним Інститутом земної кори ЛГУ, були зібрані численні залишки палеоогрганізмів, в тім числі і трилобіти, з всіх горизонтів, ордовіку, силуру і нижнього девону Поділля (опорний розріз нижньопалеозойських відкладів на Дністрі). Подальше вивчення трилобітів цих відкладів здійснював Л. І. Костантиненко.
Район дослідження. Примежові відклади силуру і девону Волино-Поділля.
Мета. Вивчити особливості розподілу фауни трилобітів силуру і нижнього девону Поділля і світу.
Завдання. 1) Вивчення морфології трилобітів верхнього силуру Волино-Поділля та світу.
2) Вивчення морфології трилобітів нижнього девону Волино-Поділля та світу.
3) Таксономічне визначення знайдених залишків трилобітів
Курсова робота виконана на кафедрі історичної геології та палеонтології під керівництвом доц., канд. геол.-мін. наук Іваніни А. В.
1. Історія визначення межі між силуром і девоном
Встановлення меж між системами, в тому числі між силуром і девоном, являються собою велику і важку задачу міжнародного характеру. Вирішити її можна тільки зусиллями вчених всіх країн світу, які у цьому зацікавлені.
Великий внесок в цю роботу зробили геологи колишньої Чехословаччини на чолі ї Йозефом Свобода. Досить цікаві і важливі дані по ФРН (проф. Г.К. Ербен) і Бельгії (проф. М. Леконт). Виключне значення мають роботи, проведені в Канаді (д-р Д. Мак-Ларен).
Проведення висновків цих досліджень було зроблено на міжнародних симпозіумах по межі та стратиграфії силуру і девону в Празі в 1958 р., в Бонне-Брюселі в 1960 р. і, спеціально по девонській системі, в Калгарі в 1967 р.
В колишньому СРСР силур і девон розповсюдженні на дуже великих площах і дуже різноманітні. Обговорення цих результатів, їх критичний розгляд був завданням Симпозіуму, проведеного в колишньому СРСР в 1968 р. У ньому брали участь геологи із 22 країн. Серед них було дуже багато видатних спеціалістів в цій області. Це надало роботі Симпозіуму більше значення. Роботи, що друкувалися у його працях, відображають останні досягнення у вивченні силуру і девону.
В більшості статей викладається зміст доповідей, зроблених на конференціях в Ленінграді, але декілька статей включають у себе результати жвавих і змістовних дискусій під час екскурсій і після них у Львові.
Під час цих дискусій були висловлені дуже різноманітні думки про положення границі між силуром і девоном. Більшість учасників висловлювалися за проведення межі в основі шарів з Monograptus uniformis під впливом видатних успіхів, досягнутих у вивченні граптолітів. Але ця думка ні в якому разі не була єдиною.
Англійські геологи і палеонтологи вказували, що оскільки силур і девон були встановленні в Англії, то англійський матеріал є вирішальним. Цей матеріал на даний час знаходиться в стадії обробки і вивчення. Тому вони просили не виносити завершальних рішень, що було і прийнято.
Остаточні рішення були перенесенні на Міжнародний геологічний конгрес 1972 р. в Канаді.
Д. В. Наливкін вважав, що для того, щоб встановлювати положення будь-якої межі між системами по розвитку однієї групи організмів, навіть такої розповсюдженої, як граптоліти, неприпустимо. Тільки кореляція розвитку всіх основних груп фауни і флори дасть матеріал, якого буде достатньо для обґрунтування границі. Особливо важливий розвиток мікроорганізмів, форамініфер, конодонтів, остракод і спор. Розвиток граптолітів повинен бути зіставлений з розвитком брахіоподів, трилобітів, ругоз, табулят, цефалоподи, можливо, пелеципод і гастропод і, у будь-якому випадку, мікроорганізмів і вапнистих водоростей.
На Симпозіумі неодноразово піднімалося питання про стратотип. Більшість учасників вважало, що для характеристики межі буде достатньо одного-двох стратотипів. Ця думка є помилковою. Для характеристики межі необхідна кореляційна таблиця із зіставленням багатьох стратотипів. Перший стратотип повинен бути для межі, що проходить серед континентальних відкладів, у тому числі древнього червоного пісковику. Другий буде для лагунних, гіпсових і соленосних відкладів. Третій – для рифових вапняків, четвертий – для шаруватих, уламкових вапняків, п’ятий – для сланцевих товщ і шостий – для піщано-сланцевих теригенних морських товщ. Напевно, знадобляться і інші. Таблиця, у якій вони будуть зіставлені, дасть той стратотиповий матеріал, який з достатньою повнотою охарактеризує будову пограничної зони. Один стратотип повинен бути замінений таблицею стратотипів.
Серед учасників Симпозіуму було багато геологів-палеонтологів, до яких і відноситься Д.В. Наливкін. Для них «палеонтологія – це все», і тільки палеонтологічних даних для проведення межі достатньо. Насправді, межа між системами – це межа між періодами історії Землі, а в історію Землі входить не тільки історія органічного світу. Навпаки, історія Землі і її періодизація ні в якому разі не визначаються однією історією органічного світу. Біосфера по порівнянню з літосферою, гідросферою і атмосферою – абсолютно другорядна річ. Не біосфера визначає історію трьох інших сфер, навпаки, її історія повністю залежить від історії інших сфер; це зрозуміло кожному.
Періодизація історії Землі – це перш за все періодизація часу. Час може бути виражений не тільки абсолютним часом. Про абсолютний вік на Симпозіумі не було ні слова.
Учасники симпозіуму були у захваті дійсно чудовими розрізами на Поділлі, вздовж р. Дністер. В тих розрізах найбільш різка зміна в історії Землі співпадає зі зміною морських вапняків і глин наземними червоноколірними пісковиками. Ця зміна спостерігається на величезних площах, цілих материках. Вона, на жаль, не притягнула достатньої уваги учасників Симпозіуму.
Ці два приклади показують, що велика і плодотворна робота Симпозіуму була в основному обмежена вивченням змін органічного світу – біостратиграфією, але ці зміни вони вивчали без будь-якої уваги до тих причин, які їх викликали. Таке вивчення не може бути досконалим.
Важливими досягненнями, котрі створили нову основу для вирішення питання про межу силуру і девону наступні.
1. Повністю відпало, що панувало в 60-х роках ХХ ст., уявлення про специфічне приурочення монограптид до силуру. Епоха розвитку граптолітів охоплює не тільки пізній лудлов, але і майже весь ранній девон, до того ж граптоліти зберігають колишню роль у зональній стратиграфії. Вище наймолодшої британської зони Saetograptus leintwardinensis тепер встановлюються ще до 15 зон (включаючи нижній емс), із котрих верхні 5-6 зон є свідомо девонськими і вже з успіхом використовуються навіть для міжконтинентальної стратиграфічної кореляції ( колишньої Чехословаччини, Марокко, Алжир, Австралія, Малайя, США, Канада; в СРСР – Поділля, Урал, Казахстан, Середня Азія).
2. Порівняно вивчення граптолітів, конодонтів, тентакулітів, трилобітів, остракод і ряду інших бентосних груп фауни різнофаціальних морських відкладів дозволило встановити, що підошва жединського ярусу Рейнсько-Арденського масиву близько відповідає підошві лохківського ярусу Баррандієна, бостовського ярусу Польщі, тиверського яруса Поділля і співпадає з основою таких важливих зон, як зона Monograptus uniformis, зона Icridus woschmidti, зона Warburgella rugulosa, кореляційна цінність яких безсумнівно встановлюється через проміжкові розрізи. Досягнуті на цій основі успіхи в стратиграфічній кореляції припадають зоні M. Uniformis s. I. Особливо велике значення.
3. Суттєво уточнилося стратиграфічне положення решток іхтіофауни в морських і перехідних розрізах Прибалтики і Поділля, що відкрило нові перспективи кореляції з фаціями Олд-Ред Великобританії (даунтонська, дитонська і бретонська серії). Зокрема, встановлено, що зона Thelodus parvidens ставить в близьке стратиграфічне положення нижній даутон (лудловський костеносний шар), минявські шари Литви, що належать уже до своєї нижньої частини до пржидольської зони Pristiograptus ultimus і горизонт Oxecaape Естонії; за іншими даними (остракоди, конодонти зони S. eosteinhornensis), до даунтону відноситься і горизонт Каугатума. Відкриття нижньодитонських Traquairaspis в тильженській світі примушує вважати ще даунтонськими і юраські шари Литви і Латвії з характерним «бейрихіїдовим» комплексом остракод, що нижче залягають.
4. В результаті проведених досліджень традиційна впевненість, що покрівля лудловського ярусу співпадає з підошвою жединського ярусу. Інше кажучи, що даунтон стратиграфічно еквівалентний нижньому жедину. Більше того, в стратиграфічній шкалі морських відкладів силуру-девону все більш ясно вимальовується пропуск, який повинен зайняти особливий стратиграфічний підрозділ, що відповідає постлудлову, але прежедину.
Верхня межа розглянутого стратиграфічного підрозділу в морських фаціях чітко визначається підошвою зон Monograptus uniformis s. I. та Icriodus woschmidti. На Поділлі вона відповідає основі тиверського ярусу, до якого О.І. Никифорова (1968) відносить і шари Тайна як нижній підрозділ борщівського горизонту.
5. Представляється, що питання про вік скальських-пржидольських відкладів та їхнє місце в стратиграфічній шкалі може вирішитися однозначно. Палеонтологічні дослідження морських безхребетних (граптоліти, брахіоподи, трилобіти, остракоди, корали, конодонти), проведені багато численними спеціалістами в різних країнах, безсумнівно, що у межах досліджуваного відрізку постлудловської історії продовжують дуже повно розвиватися типово силурійські групи фауни. Більш помітно обновляється фауна хребетних, що вторгається в морські серії постлудлова, але ця еволюційна гетерохронія зазвичай не береться до уваги при розробці еталонних стратиграфічних шкал морських відкладів.
Незалежно від таксономічного рангу цього стратиграфічного під-розділу сам факт його виявлення в морських пізньосилурійських відкладів – значне досягнення стратиграфії. Все-таки залишається невиясненим дуже важ-ливе питання про його місце стратотипової послідовності шарів Уіткліфф – даунтоська серія. Вирішенням цього питання пов’язано з визначенням межі силуру і девону в стратотипі Великобританії.
Таким чином, важлива межа зон P. transgrediens, M. uniformis, S. eosteinhornensis, I. woschmidti, настільки чітко зараз спостерігається в морських відкладах континентальної Європи і співпадає з основою жединського ярусу Рейнсько-Арденського масиву, але не може бути поки що точно вказана в стратотипових розрізах Уельса. А у зв’язку з тим тут не може бути точно виділений і об’єм верхнього підрозділу силурійської системи.
6. Результати багато численних досліджень 60-х років і думки абсолютної більшості членів спеціального міжнародного комітету, що працював під керівництвом проф. Ербена, дозволяє зробити висновок, що незалежно від того як вирішиться питання про межу силуру і девону в специфічних фаціях Великобританії, мають формальні та наукові основи прийняття рішення про нижню межу девонської системи, керуючись досягнутим рівнем біостратиграфічного вивчення жединського ярусу в стратотиповому районі і сучасними здобутками в області кореляції його морських аналогів у різних країнах світу.
У колишньому СРСР ця межа відповідає підошві тиверу, який, можливо, варто розглядати до богемської схеми як нижній підярус лохківського ярусу, який відповідає нижньому жедину або його більшій частині. Стратиграфічне положення нижньої границі добре визначаються борщівському горизонті Поділля (починаючи з шарів Тайна) M. Uniformis angustidens, I. woschmidti і багато численних видів інших груп, важливих в кореляційному відношенні. В Південній Прибалтиці на рівень цієї межі залягає тильжеська світа з Traquairaspis; в Уралі – середня (= сарайська) підсвіта петропавловської світи з Pachyfavosites kozlowskii; в Центральному Казахстані – караєспинський горизонт з Howellella mercuri, Pachyfavosites kozlowskii, Monograptus ex gr. uniformis; в Середній Азії – кунжакський горизонт (еквівалент лохкова) з Septatrypa megaera, коралами, криноїдеями, тентакулітами, монограптидами, включно з M. hercynicus; на Салаірі – томчумішський горизонт з Pachyfavosites kozlovskii, Howellella angustiplicata та своєрідним комплексом остракод; на північному сході колишнього СРСР – нелюдимська світа Favosites socialis, Protathyris praecursor та іншими видами різних груп, ще недостатньо вивчених.
Допустимі багатьма дослідниками (у тому числі і англійськими) відповідність підошви «Old Red System» підошві зони Pristiograptus ultimus, навряд чи може бути переконливою основою для зберігання status quo у відношенні цієї межі. У такому випадку довелось би вважати девонськими всі пржидольські шари, значну частину скальського горизонту, горизонт Oxecaape, ісфаринський горизонт і багато інших, силурійський вік яких ніколи не піддавався сумнівам. Неформальний підхід до вирішення цієї важкої проблеми, зв’язані історією і традиціями, повинен би швидше допустити відношення даунтона (у сенсі Аллена і Тарло, 1963) або більшої його частини до силурійської системи. Встановлення страто типів всіх ярусів девонської системи за межами Девоншира, Корнуелла і Уельса було цілком раціональним і слугує зараз виправданим прецедентом у відношенні межі силуру і девону і їх пограничних стратиграфічних підрозділів. Однак без вирішення цього питання на міжнародному рівні залишаться серйозні утруднення в області номенклатури.
2. Палеонтологічна характеристика відкладів верхнього силуру головних стратотипових розрізів світу і Волино-Поділля
В розрізі нижньопалеозойських відкладів Поділля встановлено 108 видів трилобітів, що належать 42 родам 17 родин. Із всього числа видів встановлено у силурі – 76. Вони належать 27 родам 14 родин. До верхнього силуру належить 13 видів (рис.1.). Із нижнього девону відомі 10 видів трилобітів – представників 6 родів 3 родин. Вивчення решток трилобітів і їх стратиграфічного поширення в розрізі силуру Поділля дало можливість уточнити палеонтологічну характеристику місцевих підрозділів (світ, підсвіт) підтвердити обґрунтованість виділення підрозділів регіональної схеми (горизонтів), визначити об’єми і межі ярусів (надгоризонтів) схеми силуру Поділля, а також вперше розробити зональну шкалу силуру Поділля по трилобітах (рис.2.).
Верхній відділ
Пржидольський ярус
Він представлений відкладами рукшинської серії (за винятком худиківецької світи). Нижня межа його проводиться по зміні улічського комплексу трилобітів скальським. Верхня межа ярусу (і нижня девону) проходить по підошві худиківецької світи по появі трилобітів Warburgella rugulosa, Acastella hebertiі граптолітів Monograptus uniformis. Скальський комплекс трилобітів нечисленний, включає види родів Acaste, Calymene, Proetus, Warburgella, Acastella, Tetinia. В нижній частині розрізу, пригородська і варницька світи, зустрінуті Acaste podolica, Calymene scalensis, Proetus scalicus по поширенню котрих виділені трилобітові зони Acaste podolica (= граптолітовій зоні ultimus-parultimus) і Calymene skalensis (= граптолітовій зоні lochkovensis і нижній частині граптолітової зони tranagrediens-bouceki). Відклади трубчинської (верхня частина) і звенигородської світ вміщують рештки Calymene dnestroviana, Acaste dayiana, Proetus dnestrovianus, P. volkovcyanus, P. scalicus, Tetinia minuta, які вище в розрізі відсутні. По присутності Calymene dnestroviana, Acaste dayiana, Tetinia minuta вони співставляються з горизонтом Oxecaape Естонії, з подляськими верствами Польщі і пржидольськими верствами Чехії, відповідають трилобітовій зоні Calymene dnestroviana (= верхній частині граптолітової зони transgrediens-bouceki). Худиківецька світа віднесена до нижнього девону. Вона відповідає трилобітовій зоні Warburgella rugulosa(= граптолітовій зоні uniformis).
В розрізі силуру Поділля трилобіти було поширенні від мілководної до зони материкового схилу включно, але чисельність їх і видовий склад в кожній із зон були різні. Найбільш поширеними були трилобіти на ділянках дна з м’яким карбонатним мулом, збагаченим органогенним детритом (зони глибокого шельфу і схилу).
Угр.упування Acaste podolica малочисленне за складом. Ха- рактерні Acaste podolіca, Calymene cf.tentaculata, Proetus cf.kure- ssaarensis. - Приурочено до мілководних відкладів ісаковецької під-світи і нижньої частини варницькоі світи. Існувало в умовах зони за-критого шельфу та барової зони на твердому карбонатному субстраті. Склад відкладів дозволяв відносити це угрупування до другого бентос-ного комплексу.
Угр.упування Proetus scalicus приурочено до відкладів варни-цької світи, нечисленне. Характерні Proetus scalicus, Calymene skalensis i поодинокі Proetus cf.pulcher. Існувало в умовах спокійного середовища на мулистому субстраті. Характерно для нижньої части-ни літоральної і верхньої частини субліторальної зон області другого бентосного комплекса.
Угр.упування Calymene dneatroviana більш численне і різноманітне за складом. Відноситься до типу угрупувань Acaste dayiana /Мянніль, 1982/. Приурочено до відкладів трубчинської і звенигородської світ. Разом з Calymene dnestroviana зустрінуті Proetus volkovcyanus, P.dnestrovianus, Асаste dayiana, Calymene skalensis, Proetus scalicus, P.rashkoviensis, Tetinia minuta, Warburgella vol- kovcyana, Acastella sp. Угрупування існувало в ареалі третього бентосного комплексу в умовах відкритого шельфу на мулистих грунтах з тенденцією поглиблення басейну і швидкої зміни фацій від шельфу до схилу.
Систематичний склад описаних трилобітів наступний:
Клас Trilobita
Загін Polymera
Родина Proetidae Hawle et Corda, 1847
Підродина Proetinae Hawle et Corda, 1847
Рід Proetus Steininger, 1831
Proetus cf. kuressaarensis Munnil, 1981
P. scalicus Balashova, 1968
P. rashkoviensis sp.n.
P. dnestrovianus Balashova, 1968
P. volkovcyanus Balashova, 1968
Родина Warburgella Reed, 1931
Підродина Warburgella (Warburgella) Redd, 1931
Warburgella (Warburgella) volkovcyana Balashova sp. nov
Рід Tetinia Clupac, 1971
Tetinia minuta (Pribyl et Vanek, 1962)
Родина Calymenidae Burmeister, 1843
Рід Calumene Brongniart, 1822
Calymene cf. tentaculata (Schlotheim, 1820)
C. skalensis Munnil, 1983
C. dnestroviana Balashova, 1968
Загін Calmonidae Dello, 1935
Підзагін Acastinae Dello, 1935
Рід Acaste Goldfuss, 1843
Acaste dayiana R. et E. Richter, 1954
Acaste podolica Balashova, sp. nov.
|
Види |
Система |
Силурійська |
||
|
Ярус |
Пржидольський |
|||
|
Серія |
Рукшинська |
|||
|
Світа |
Пригородокська |
Варницька |
Трубчинська |
Звенигородська |
|
Proetus cf. kuresaarensis Munn |
……………. |
-- |
||
|
Acaste podolica Bol. |
……………. |
-- |
||
|
Calymene cf. tentaculata Schlot. |
……………. |
--- |
||
|
Proetus cf.pulcher Niezk. |
------……-- |
|||
|
P. scalicus Bal. |
------- |
--------------- |
----------- |
|
|
Calymene skalensis Munn. |
------- |
--------------- |
---------------- |
|
|
C. dnestroviana Bal. |
-------- |
---------------- |
||
|
Acaste daylana R. et E. Richt. |
-------- |
---------------- |
||
|
Proetus dnestrovianus Bal. |
---------------- |
|||
|
P. volkovcyanus Bal. |
---------------- |
|||
|
P. rashkoviensis sp.n |
--------- |
|||
|
Warburgella volkovcyana Bal. |
----- |
|||
|
Tetina minuta (Pr. et Van.) |
--- |
Рис.1. Стратиграфічне поширення видів трилобітів у розрізі верхнього силуру Поділля.
|
система |
відділ |
Регіоярус (н/горизонт) |
горизонт |
Трилобіти (Константиненко, 2003) |
|
|
Силурійська |
S4 |
скальський |
склавинський |
Calymene dnestrowiana |
|
|
Calymene skalensis |
|||||
|
станавський |
|||||
|
Calymene tentaculata |
|||||
|
S3 |
улицький |
метонський |
Calymene spectabilis |
|
|
|
тагринський |
|
||||
|
S2 |
тиритський |
конівський |
|||
|
Proetus conspersus |
|||||
|
неврідський |
|||||
|
Calymene mukschaensis |
|||||
|
S1 |
китайгородський |
алізонський |
|||
|
Calymene blumenbachi |
|||||
|
фурманівський |
|
||||
|
болотинський |
|||||
|
Calymene frontosa |
Рис.2 Стратиграфічна схема силурійських відкладів України по трилобітах
3. Палеонтологічна характеристика відкладів нижнього девону головних стратотипових розрізів світу і Волино-Поділля
В розрізі нижньопалеозойських відкладів Поділля встановлено 108 видів трилобітів, що належать 42 родам 17 родин. Із всього числа видів встановлено у силурі – 76. Вони належать 27 родам 14 родин. Із нижнього девону відомі 10 видів трилобітів – представників 6 родів 3 родин (рис.3). Вивчення решток трилобітів і їх стратиграфічного поширення в розрізі девону Поділля дало можливість уточнити палеонтологічну характеристику місцевих підрозділів (світ, підсвіт) підтвердити обґрунтованість виділення підрозділів регіональної схеми (горизонтів), визначити об’єми і межі ярусів (надгоризонтів) схеми девону Поділля, а також вперше розробити зональну шкалу девону Поділля по трилобітах (рис.4).
Тиверський надгоризонт
Борщівський горизонт
Тайнівські шари. В шарах Тайна, розкритих вздовж р. Дністер, зустрічаються рештки фауни: табуляти, ругози, пелециподи, наутилоідеї, моховатки, брахіоподи, остракоди, трилобіти – Encrinurus subvariolaris concomitans Prib. et Vanek, Warburgella heberti elsana Richt. R. E., W. rugulosa Alth, W. rugulosa rugosa Bouc., Acastella cf. prima Tom., A. tiro Richt. R. E., A. heberti elsana Richt. R. E.; конодонти, граптоліти, криноідеї.
Митківські шари. Загальний склад фауни митківських шарів такий: табуляти, ругози, пелециподи, наутілоідеї, моховатки, брахіоподи, трилобіти - Warburgella rugulosa Alth, W. rugulosa rugosa Bouc., Acastella tiro Richt. R. E., A. heberti elsana Richt. R. E., Homolanotus roemeri Kon., Proetus bohemicus Barrande; остракоди, криноідеї, граптоліти, риби. Трилобіти митківських шарів - Warburgella rugulosa rugosa Alth., W. rugulosa rugosa Bouc, Acastella tiro Richt. R. E., Homolanotus roemeri Kon. – разом з граптолітами – Monograptus ex gr. uniformis Prib. – і конодонтами Icriodus woschmidti Ziegl. – виявились формами, широко відомими і добре вивченими на даний час в ФРГ і ГДР, Марокко і Чехословакії, що дозволяє вважати цю частину борщівського горизонту аналогічною нижньому жедину Бельгії, шарам Huinghauser Західної Німеччини і нижньому лохкову колишньої Чехословаччини.
Богданівські шари. Склад фауни богданівських шарів з порівнянням з митківськими бідніший. На їх рівні зникають роди і види, що раніше зустрічалися табулят і ругоз, повністю відсутні рештки трилобітів. Присутні пелециподи, наутилоідеї, тентакуліти, моховатки, брахіоподи, остракоди, криноідеї, конодонти.
Чортківський горизонт
Загальний склад чортківського горизонту наступний: табуляти, пелециподи, моховатки, брахіоподи, наутилоідеї, остракоди, тентакуліти, криноідеї, риби, конодонти. Трилобіти відсутні.
Іванівський горизонт
Рештки фауни в іванівському горизонті залишаються такими ж як і в чортківському горизонті, крім остракод, які тут настільки різко обновляють свій склад, що служать приводом для виділення самостійності іванівського горизонту. Трилобітів немає.
Систематичний склад описаних трилобітів наступний:
Клас Trilobita
Загін Polymera
Рід Warburgella Reed, 1931
Підрід Warburgella (Warburgella) Reed, 1931
Вид Warburgella (Podolites) rugulosa (Alth, 1874)
Вид W. (Podolites) rugosa rugosa (Boucek, 1934)
Загін Homalonotidae Chapman, 1890
Рід Homanolotus Koenig, 1825
Вид Homanolotus (Digonus) roemeri (Koninck, 1867)
Загін Dalmanitidae Vogdes, 1890
Підзагін Acastavinae Struve, 1958
Рід Acastella Reed, 1925
Вид Acastella heberti elsana R. et E. Richter, 1954
Вид Acastella tiro R. et E. Richter, 1954
|
Види |
Система |
Девонська |
|
ярус |
жедин |
|
|
підярус |
Нижній жедин |
|
|
надгоризонт |
тиверський |
|
|
горизонт |
борщівський |
|
|
шари |
тайнівські |
митківські |
|
Encrinurus subvariolaris concomitans Prib. et Vanek |
-------------- |
|
|
Warburgella heberti elsana Richt. R. E. |
-------------- |
|
|
W. rugulosa Alth |
-------------- |
-------------- |
|
W. rugulosa rugosa Bouc. |
-------------- |
-------------- |
|
Acastella cf. prima Tom., |
-------------- |
|
|
A. tiro Richt. R. E., |
-------------- |
-------------- |
|
A. heberti elsana Richt. R. E.; |
-------------- |
-------------- |
|
Homolanotus roemeri Kon., |
-------------- |
|
|
Proetus bohemicus Barrande |
-------------- |
Рис.3. Стратиграфічне поширення видів трилобітів у розрізі нижнього девону Поділля.
|
Система |
Яруси, підяруси, надгоризонти |
Горизонти |
трилобіти |
||
|
Девон |
Жедин |
Верхній жедин |
Дністровська серія |
- |
|
|
Нижній жедин |
Тиверський надгоризонт |
Іванівський |
- |
||
|
Чортківський |
- |
||||
|
Борщівський |
7 |
||||
|
Силур |
Надлудловський |
Скальський |
8 |
||
|
Лудловський |
|||||
|
Малиновецький |
4 |
||||
|
Венлокський |
Устєвський |
- |
|||
|
Мукшинський |
4 |
||||
|
Ландоверський ? |
Китайгородський |
6 |
|||
|
Ордовік |
Ашгілський |
Молодовський |
2 |
Рис.4. Кількісний розподіл родів трилобітів в розрізі силуру і нижнього девону Поділля
4. Морфологічний опис деяких видів трилобітів примежових відкладах силуру і девону Волино-Поділля.
Трилобіти родин Proetidae, Acaste і Warburgella, представники якого поширені по всьому розрізу верхнього силуру Поділля, мають важливе значення для стратиграфії.
Загін Polymera Jaekel, 1909
Родина Proetidae Salter, 1864
Рід Proetus Steininger, 1831
Proetus (Proetus) volkovcyanus Balashova, nov
Табл. 1, фіг. 1
Опис. Кранідій слабо випуклий. Передня крайова кайма ширша потиличного кільця, струмениста. Глабель слабо і рівномірно випукла, майже циліндрична, напереді притуплена, відділена виразною борозною від крайової кайми. Глабельні борозни не виражені. Потилична борозна широка, трішки випукла. На кінцях спостерігаються потиличні лопати. Спинні борозни виразні. Очні кришки широкі, дотикаються до спинних борозен, починаються трохи спереду середини довжини глабелі і не досягає потиличної борозни. Поверхня кранідія покрита дуже маленькими горбками, що не дотикаються один до одного. Пігідій напівкруглий слабко випуклий. Рахіс досить високий, конічний, майже плоский, слабко обмежений розпливчастою борозною, лімба. Кільця трішки випуклі, розділені широкими спереду, звужуючими назад борознами. Їх 9. Спинні борозни виразні. Плеври полого дугоподібно зігнуті. Бокових ребер 7 пар. Є місце для 8-ї пари. Вони плоскі, широкі, на зовнішній частині сильно відхиляються назад й трохи спостерігаються на лімбі. Міжплевральні борозни ширші плевральних. На усій поверхні, за виключенням борозен, спостерігаються маленькі горбики, а на лімбі на ребрах – по одному струмені
Розповсюдження. Поділля, скальський горизонт
Родина CalmonidaeDello, 1935
Підродина Acastinae Dello, 1935
Рід Acaste Goldfuss, 1843
Acaste dayiana R. et E. Richter, 1954
Табл.1.фіг. 2.
Опис. Пігідій сильно випуклий, з вузьким, плоским лімбом, що продовжує схил плевр. На ядрах він обмежений невеликим перегином, слабко випуклий. Рахіс випуклий, з округленим поперечним перерізом, конічний, трішки не досягає лімбу. Не менше 7-8 виразних кілець, розділених різкими борознами. Кінець його не розчленований, на ньому вміщуються 1-2 ребра. Шість пар випуклих ребер, трішки витягнутих вперед. Передні п’ять ребер продовжують кільця. Плевральні борозни дуже вузькі, найкраще виражені біля спинних борозен і на внутрішній частині плевр, спостерігаються до зовнішнього кінця ребер, які не досягають кінця пігідія.
Розташування. Поділля, скальський горизонт
Рід Warburgella Reed, 1931
Підрід Warburgella (Warburgella) Reed, 1931
Warburgella (Warburgella) volkovcyana Balashova sp. nov
Табл.1. фіг. 3.
Опис. Головний щит напівкруглий, плоский. Передня крайова кайма горизонтальна, плоска, трішки ширша розташованої на заді її плоскодонної борозни без валика. Ззаду ця борозна обмежена ниткоподібним тропідієм, що проходить одразу спереду передглабельної борозни і до лицевих швів, що згладжуються. Глабель досить сильно розширяється назад, дуже слабко випукла. Наявні дві пари глабельних борозен. Передні простежуються менш ніж на 1/3 ширини глабелі. Задні борозни глибокі, сильніше передніх відхилені назад, спереду досягають спинних борозен, ззаду – потиличної борозни. Базальні лопати помірно трикутні, випуклі. Лопатки передніх і задніх пар на ядрах виразно обмежені, випуклі. Потиличне кільце майже плоске з виразною серединною туберкулою. Потиличні лопатки слабо випуклі. Очі спостерігаються від передніх борозен до заднього кінця базальної лопатки. Передні гілки лицевого шва сильно розходяться в сторони, але на передній крайовій каймі відхиляється всередину. Спинні борозни виразні на всьому простяганні і зливаються з передглабельною борозною. Вільні щоки слабо і рівномірно випуклі, без тропідія. Ширина бокової і задньої крайової борозни однакова, але задня глибше; вони зливаються і продовжуються вздовж щокового шипа. Бокова крайова кайма плоска, задня – слабо випукла. По усій поверхні щита спостерігаються неправильної форми, різних розмірів тісно сидячі горбики, котрі на глабелі розташовані у ряди, вигнуті уперед. Вздовж внутрішнього краю щита і шипів тягнуться 1-2 подовжні цівки.
Пігідій напівкруглий, плоский. Рахіс конічний, трішки випуклий, не досягає неглибокої, широкої борозни, що обмежує слабо випуклий або плоский лімб. Кілець до 11. Вони плоскі. На поверхні пігідія горбки капле видні, різного розміру, тісні. На середній частині кілець вони розташовані у ряди, дугоподібно витягнуті вперед. Ряд горбочків, що ідуть вздовж заднього краю кільця, особливо виразний, що робить межу між кільцями різкою. На схилах рахісу горбочки розташовані хаотично. Зазвичай на передньому краю пігідія спостерігається ряд горбочків вздовж заднього краю передньої гілки бокових ребер. На іншій поверхні ребер і лімбі вони розташовані хаотично. Ширина дублери головного щита і пігідія рівна ширині лімба, поверхня її покрита цівками.
Розповсюдження. Верхня частина скальського горизонту. Поділля.
Трилобіти родин Acastella, Homalonotus і Warburgella, представники якого поширені по всьому розрізу нижнього девону Поділля, мають важливе значення для стратиграфії.
Рід Warburgella Reed, 1931
Підрід Warburgella (Warburgella) Reed, 1931
Warburgella (Podolites) rugulosa (Alth, 1874)
Табл.2. фіг.1.
Опис. Головний щит напівкруглий, трохи випуклий. Передня крайова кайма трішки при піднята, дуже слабко випукла (майже горизонтальна), трохи ширша передглабельного поля. На ній є дві борозни. Передня дуже слабо виражена, згладжується, не доходячи до лицевих швів. Задня різка, що розширяється до лицевих швів, що обмежує спереду виразний валик (тропідій), що лежить спереду глибокої, вузької передглабельної борозни. Цей валик спостерігається до задньої крайової борозни.
Глабель трохи розширяється назад, слабо випукла, найбільше на рівні очей. Передній схил її дуже пологий, задній – крутіший. Базальні лопатки трикутно яйцевидні, видовжені, випуклі.
Довжина кранідія великих розмірів 6 мм, ширина кранідія на задньому краю передньої кайми 4 мм. Довжина глабелі 3,8 мм, ширина глабелі спереду очей 2,6 мм, ширина глабелі ззаду 3 мм. Гіпостома з ввігнутим заднім краєм. Середнє тіло її спереду випукле, розширяється і понижується назад. Задня лопатка дугоподібна зігнута назад, обмежена спереду розпливчатою, ззаду – виразною борозною. Макули не помітні.
Тулуб із 8 сегментів. Рахіс випуклий, вже плевр. Діагональні борозни виразні. Пігідій напівкруглий. Рахіс випуклий, циліндричний, високий, досягає крайової борозни.
Розповсюдження. Поділля, борщівський горизонт; Польща, бостовські шари; ФРН, нижній жедин.
Родина Homalonotidae Chapman, 1890
Рід Homalonotus Koenig, 1825
Homalonotus (Digonus) roemeri (Koninck, 1867)
Табл.2.фіг.2.
Опис. Головний щит трикутний. Передній край його на середині загострений. Глабель звужується вперед, слабо випукла, обмежена розпливчастими спинними борознами. Передглабельне слабо ввігнуте. Потиличне кільце вузьке, слабо випукле. Крайові борозни виразніші потиличних. Задня крайова кайма слабо випукла, розширяється до щокових шипів. Очі великі, відділені від вільних щік невиразною борозною. Передні гілки лицевого шва зближуються, на передглабельному полі круто повертаються і зливаються разом. Задні гілки перетинають бічний край поблизу щокового кута. Тулуб сильно випуклий. Рахіс майже у два рази ширший плевр. Кільця випуклі з горбистою поверхнею. Плеври із закрученими кінцями. Задня частина плевр горбиста. Пігідій трикутний, випуклий. Бічні краї його плавно переходять в короткий, тонкий шип. Задній схил рахісу порівняно крутий. Кілець 11, ребер 10 пар, вони слабо випуклі, розмежовані вузькими борознами. На поверхні свого спинного панцера спостерігаються пори, що ведуть у канали чуттєвих щетинок.
Розташування. Поділля, борщівський горизонт, верхня частина; Зх. Європа, нижній жедин.
Висновки
Трилобіти є важливою для біостратиграфії верхнього силуру та нижнього девону групою організмів. Рівні найбільш істотних змін видового і родового складу їх приурочені до меж стратиграфічних підрозділів рангу горизонтів і надгоризонтів.
Аналіз поширення решток трилобітів в розрізі девону та силуру Поділля дозволив виділити ряд угрупувань, що дає можливість корелювати вміщуючи їх відклади як в регіоні, так і за його межами.
Список використаної літератури:
1. Вебер В.Н. 1951. Верхнесилурийские трилобити СССР. М. Госгеолтехздат.
2. Граница силура и девона и биостратиграфия силура Сост. Л. А. Рейхерт и др. – Ленинград: «Наука», 1968, с. 282.
3. Л.І. Константиненко. Атореферат дисертації на здобуття вченого ступеня кандидата геолого-мінералогічних наук. – Київ, 1993, с. 22.
4. Опорний разрез силура и нижнего девона Подолии . Изд-во «Наука», Л., Ленингр. отд., 1972, I – 262.
5. Палеонтологічний збірник. 2005. № 37. 143 с.
6. Палеонтологічний збірник. 2006. № 38. 143 с.
7. Палеонтологічний збірник. 2007. № 39. 143 с.
8. Палеонтологічний збірник. 2008. № 40. 143 с.
9. Палеонтологічний збірник. 2009. № 41. 143 с.
10. Путеводитель краткого геологического маршрута по разрезам верхне-протерозойских и палеозойских отложений Подольского Приднестровья / Сост. И. Б. Вишняков, Г. М. Помяновская, Ю. Н. Сеньковский, А. В. Хижняков. – Львов: Вища школа, 1981, с. 40.
11. Силурийсько-девонская фауна Подолии З. Г. Балашов и др. – Ленинград:, 1968, с. 123.
12. Силур Подолии: Путеводитель экскурсии – The Silurian of Podolia: A guide to the excursion Сост. П. Д. Цегельнюк и др. – Киев : Наук. думка, 1983, с. 224.
13. Трилобитовые сообщества силура Подолии. – В сб: «Биостратиграфия, палеонтология осадочного чехла Украины». – Наукова думка, Киев, 1987, с. 27-30
14. Richter r. et E. 1954. Die trilobiten des Elbe Sattels und zu vergleichnde Arten.
Додаток 1. Табл. 1.
|
|
фіг.1 Фрагмент пігідія трилобіта роду Proetus (Proe-tus) volkovcyanus Bala-shova, nov |
|
|
фіг.2 Фрагмент пігідія трилобіта роду Acaste dayiana R. et E. Richter, 1954 |
|
|
фіг.3 Фрагмент пі-гідія трилобіта ро-ду Warburgella (Warburgella) volkovcyana Balashova sp. nov |
Додаток 2. Табл.2
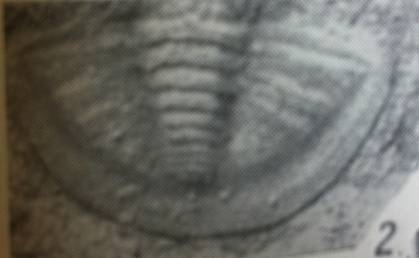
|
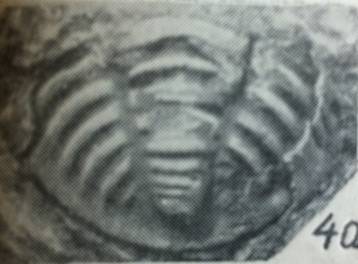
|
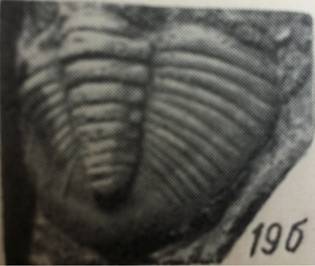
фіг.1 Фрагмент пігі-дія трилобіта роду Warburgella rugulosa Alth |
|
|
фіг.2. Фрагмент пігі-дія трилобіта роду Homalonotus (Digonus) roe-meri (Koninck, 1867) |


 (zip - application/zip)
(zip - application/zip)