Биохимия и ее задачи
Биохимия – это наука о структуре веществ, входящих в состав живого организма, их превращениях и физико-химических процессах, лежащих в основе жизнедеятельности.
Биохимия является одной из фундаментальных дисциплин медицины и биологии. Она занимается познанием живого на уровне макромолекул. Биохимия – это результат интеграции биологии и химии.
Разделы биохимии:
1. статическая (биоорганическая химия);
2. динамическая (изучает превращение веществ);
3. функциональная (изучает физико-химические процессы).
Выделяют разделы биохимии в зависимости от объекта изучения: биохимия животных, микроорганизмов, растений, человека, клиническая биохимия и т.д.
Основные задачи биохимии:
1. изучение процессов биокатализа;
2. изучение строения и функций нуклеиновых кислот;
3. изучение молекулярных механизмов наследственности;
4. изучение строения, обмена белков;
5. изучение превращения углеводов;
6. изучение процессов обмена липидов;
7. изучение роли биорегуляторов (гормоны, нейромедиаторы);
8. изучение роли витаминов и минеральных веществ.
Значение БХ для медицины:
1. необходима для понимания сущности заболевания (патогенеза), его механизма. Пр.: сахарный диабет в результате недостатка инсулина, атеросклероз – нарушение обмена липопротеинов, опухолевый рост – функционирование онкогенов;
2. необходима для диагностики заболеваний. Пр.: биохимический анализ крови, мочи. Определяется:
а) количество субстрата (уровень метаболита);
б) активность фермента;
в) количество биорегуляторов (гормонов и нейропептидов);
В диагностике заболеваний используются различные методы: радио-иммуный анализ, иммуно-ферментный анализ, введение ДНК-зондов для выявления чужеродной ДНК, дефектов ДНК, онкогенов. Также позволяет выявить предрасположенность к заболеваниям;
3. разработка новых лекарственных препаратов;
4. необходима для профилактики заболеваний. Пр.: рахит – в результате недостатка витамина D, цинга – витамина С.
Белки и их биологическая рольБелок (протеины) – protos – предшествующий всему, первичный, наиглавнейший, определяющий всё остальное.
Белки – это высокомолекулярные азотсодержащие органические вещества, состоящие из аминокислот, соединённых в цепи с помощью пептидных связей и имеющих сложную структурную организацию.
Основные отличительные признаки белков:
1. содержат азота больше, чем другие вещества (16%). Так, 1г азота содержится в 6,25г белка;
2. состоят из альфа-аминокислот L-ряда;
3. наличие пептидных связей;
4. большая молекулярная масса (от 4-5 тыс. дальтон до нескольких млн.);
5. имеют сложную структурную организацию;
6. белки составляют 25% сырой ткани и 45-50% сухой ткани.
Биологическая роль белков:
1. каталитическая (выполняют ферменты);
2. структурная, т.е. белки являются основным компонентом клеточных структур;
3. регуляторная (выполняют белки-гормоны);
4. рецепторная, т.е. рецепторы клеточных мембран имеют белковую природу;
5. транспортная – белки участвуют в транспорте липидов, токсических веществ, кислорода и т.д.;
6. опорная – выполняет белок коллаген;
7. энергетическая. Заключается в том, что при окислении 1г белка выделяется 17,6 кДж (4,1ккал) энергии;
8. сократительная – её выполняют белки актин и миозин;
9. генно-регуляторная – её выполняют белки гистоны, участвуя в регуляции репликации;
10. имуннологическая – её выполняют белки антитела;
11. гемостатическая – участвуют в свёртывании крови, препятствуют кровотечению;
12. антитоксическая, т.е. белки связывают многие токсические вещества (особенно соли тяжёлых металлов) и препятствуют развитию интоксикации в организме.
Физико-химические свойства белков:
Структура белка определяет его свойства. Существует несколько групп свойств.
I. Электрохимические свойства белков:
1. белки - амфотерные полиэлектролиты (амфолиты). Это достигается за счет наличия концевых СОО- и NH3+ групп, а также ионогенных групп боковых радикалов (ГЛУ, АСП, ЛИЗ, АРГ, ГИС)
2. буферность белков (поддержка рН среды). При физиологических значениях рН буферные свойства ограничены и обусловлены наличием кислотных и основных групп. Наибольшим буферным действием обладает гистидин, которого много в гемоглобине, за счет чего последний является мощным буфером крови;
3. наличие заряда в белковой молекуле. Обусловлено соотношением кислых и основных АК, а также ионизацией бокового радикала. Степень ионизации зависит от рН среды. Так, если среда кислая, то ионизация СООН групп заторможена и белок приобретает «+» заряд. В щелочной среде заторможена ионизация NH2 групп и белок заряжается «--».
Изоэлектрическое состояние белка наступает, когда заряд белковой молекулы равен 0, а рН среды, при котором белок находится в изоэлектрическом состоянии, называется изоэлектрической точкой (рI). Она определяется соотношением кислых и основных радикалов. У большей части белков цитоплазмы рI меньше 7, т.е. эти белки кислые; у ядерных белков больше 7, т.е. они основные.
Наличие заряда используется для разделения белков с помощью электрофореза – движения белков в электрическом поле. Наличие заряда обусловливает устойчивость в растворе. В изоэлектрическом состоянии белки наименее устойчивы и выпадают в осадок.
II. Коллоидные свойства.
Растворы белков чаще всего достаточно устойчивы. Хорошая растворимость приближает растворы белков к истинным растворам, но высокая молекулярная масса придает им свойства коллоидных систем:
1. способность рассеивать свет (опалисценция). Наблюдается помутнение при боковом освещении - эффект Тиндаля [рис. рассеивающегося луча]. Используется в световой микроскопии (нефелометрии);
2. малая скорость диффузии;
3. высокая вязкость растворов белков;
4. неспособность белков проникать через полупроницаемые мембраны (явление осмоса). На этом основан диализ – очищение белков;
5. способность белковых растворов образовывать гель. Наиболее выражено у фибриллярных белков.
III. Гидрофильные свойства.
Белки хорошо связываются водой, обусловлено наличием полярных гидрофильных групп. Вода может проникать в белок и связываться с его гидрофильными группами, вызывая его набухание. Также возможно образование гидратной оболочки. 100г белка связывают 30-35г воды.
IV. Растворимость белков.
Чем больше полярных групп содержит белок, тем больше он растворим. Глобулярные белки растворяются лучше. Растворимость белков зависит от 2-х факторов:
- наличия заряда;
- образования гидратной оболочки.
Чтобы осадить белок, необходимо ликвидировать эти 2 фактора. Осаждение белков с помощью нейтральных солей называется высаливание – обратимое осаждение. После удаления высаливающегося фактора белок сохраняет все свои свойства.
V. Денатурация.
Под действием внешних факторов нарушается высшие уровни (вторичный, третичный, четвертичный) структурной организации белков с сохранением первичной структуры. При этом белок теряет свои нативные свойства. При денатурации разрываются связи, удерживающие высшие структурные организации. Денатурацию вызывают физические и химические факторы: давление, температура, механическое воздействие, ультразвук, ионизирующее излучение, кислоты, щёлочи, органические растворители, соли тяжёлых металлов. При кратковременном воздействии денатурирующих факторов возможна ренатурация.
Классификация белков
В настоящее время насчитывается ~5 млн. белков. Их пытались классифицировать по физико-химическим свойствам, например по растворимости, плотности, форме молекул (глобулярные и фибриллярные), локализации и происхождению, АК-составу, биологической роли. Однако все эти классификации не соответствуют тем знаниям о белках, которые известны на сегодняшний день.
В основе классификации лежит химический состав белка. По этому признаку все белки делят на простые и сложные.
Простые белки – это белки, образованные только полипептидными цепями, состоящие только из АК-ных остатков.
Сложные белки имеют две части: белковая или пептидная построена из АК-ных остатков, и небелковая (простетическая) часть.
К простым белкам относят: гистоны, протамины, альбумины, глобулины, глютелины, проламины и протеноиды (склеропротеины).
К сложным белкам относят: хромопротеины, нуклеопротеины, фосфопротеины, углевод-белковые и липид-белковые комплексы.
Связь белковой части с небелковой может быть ковалентной, ионной и др.
Характеристика простых белковВ основе классификации (создана в 1908г.) лежит растворимость белков. По этому признаку выделяют:
I. гистоны и протамины, растворимые в солевых растворах. Они относятся к низкомолекулярным белкам (M<5 000 дальтон, Да). В своем составе эти белки содержат большое количество диаминомонокарбоновых кислот (аргинина и лизина). В водной среде они имеют «+» заряд, изоэлектрическая точка (ИЭТ) лежит в области pH 9-12 (щелочная среда).
Основные функции протаминов и гистонов:
1. структурная, т.е. они поддерживают пространственную организацию ДНК и входят в состав ДНП.
2. регуляторная - гистоны регулируют процесс транскрипции.
Протамины имеют меньшую молекулярную массу, чем гистоны; они содержат больше аргинина (до 85%). Происхождение протаминов: сальмин выделен из молок лососевых рыб, скумбрин – скумбрии.
II. Альбумины и глобулины. Это наиболее важные белки плазмы крови (составляют 60%). Содержание альбумина 35-50 г/л, глобулина 20-30 г/л. По форме молекулы это глобулярные белки.
Альбумины растворимы как в воде, так и в солевых растворах, имеют M 40-70 кД (килодальтон, т.е. тысяч дальтон). Особенностью АК-состава является повышенное содержание глутаминовой, аспарагиновой кислот, следовательно в водном растворе эти белки имеют «-» заряд. ИЭТ лежит в пределах pH 4,7 в кислой среде.
В организме альбумины выполняют следующие функции:
1. транспортная – переносят метаболиты, лекарственные вещества, ВЖК (высшие жирные к-ты), холестерины, гормоны, желчные пигменты, ионы Ca2+ и др.
2. поддержание онкотического давления, т.к. осмотическое давление крови на ~75-80% обеспечивается за счет альбуминов.
Глобулины плохо растворимы в воде, но хорошо в солевых растворах. Имеют большую M до 150 кД. Проявляют либо слабовыраженные кислотные, либо нейтральные свойства. ИЭТ лежит в пределах pH 6-7,3.
В норме содержание альбуминов и глобулинов лежит в определенной пропорции – альбумино-глобулиновый или белковый коэффициент: К=А/Г»1,5-2,3. Определение АГК имеет диагностическое значение, т.к. при патологиях оно меняется. Например, при инфекционных заболеваниях увеличивается содержание глобулина, АГК уменьшается; при патологии почек нарушается их фильтрационная способность, количество альбуминов уменьшается, следовательно, АГК также падает. Чтобы определить белковый коэффициент необходимо отнести содержание альбумина к содержанию глобулина.
Методы разделения (фракционирования) белков1. Высаливание – разделение на основе различной растворимости альбуминов и глобулинов. Осаждение белков обычно производят сульфатом аммония (NH4)2SO4. В насыщенном растворе этого реактива осаждаются альбумины, а в полунасыщенном – глобулины.
2. Электрофорез – разделение белков при движении в электрическом поле за счет разности их заряда.
Основные белки сыворотки крови делятся на несколько фракций. Быстрее всех движутся к аноду альбумины – это гомогенная фракция. Глобулины делятся на 4 фракции: a1, a2, b, g [рис. альбуминов и 4-х глобулинов и как они движутся к аноду].
В крови определяется общий белок. Нормальное общее содержание белка, но изменено соотношение его фракций – диспротенемия. При инфекционных заболеваниях увеличивается содержание g-глобулиновой фракции. При заболеваниях почек снижается содержание альбуминовой фракции, но увеличивается содержание a2 и b глобулинов. Также наблюдается парапротеинемия – появление патологических белков, например при некоторых онкологических заболеваниях.
III. Проламины и глютелины. Это основные растительные белки. Они не растворяются ни в водных растворах, ни в свободном этаноле, но растворяются в 65% растворе этанола. По АК-составу присутствует глутамин (до 25%), пролин (до 15%). В кукурузе содержится зеин, в ячмене – гордеин, в пшенице – глиадин.
IV. Протеиноиды (склеропротеины). Они не растворяются ни в воде, ни в солевых растворах. Это фибриллярные белки опорных тканей. Входят в состав сухожилий, костной, хрящевой тканей и др. К ним относятся: кератины – белки волос, ногтей; коллагены – белки соединительной ткани; эластины – белки, входящие в состав связок. Особенностью АК-состава является повышенное содержание аланина, глицина, пролина. Имеется оксипролин. Эти белки составляют 25-33% от всех белков в организме.
Характеристика сложных белков
Они имеют белковую и небелковую (простетическую) части. Белковую часть составляет полипептид, построенный из АК-остатков. В состав небелковой части может входить: гем, металл, остаток фосфорной кислоты, углеводы, липиды и т.д.
Хромопротеины
Для них простетическая часть окрашена (chromos – краска). К хромопротеинам относятся гемоглобин, миоглобин, каталаза, пероксидаза, ряд флавинсодержащих ферментов (сукцинатдегидрогеназа, альдегидоксидаза, ксантиноксидаза), цитохромы (гемсодержащие белки) и т.д. Велика биологическая роль этих белков – участвуют в физиологических процессах: дыхание клетки, транспорте кислорода и углекислого газа, окислительно-восстановительных процессах.
Гемоглобин. Его белковая часть представлена глобином, небелковая – гемом. Это олигомерный белок, т.е. имеет четвертичную структуру, состоящую из 4 субъединиц.
a цепи построены из 141 АК-остатка.
b цепи из 146 АК-остатков [рис. 4-х субъединиц, в каждой нарисована точка - гем].
Каждая из субъединиц связана с гемом:
 [гем]
[гем]
Основная функция гемоглобина – транспортная (кислород, углекислый газ). Также он представляет собой основную буферную систему крови (75% от всей буферной емкости крови).
Различают:
- HbO2 – оксигемоглобин (связан с молекулой O2);
- HbCO – карбоксигемоглобин;
- HbCO2 – карбгемоглобин;
- HbOH – метгемоглобин (образуется при соединении с нитросоединениями, не способен связывать кислород).
Типы гемоглобина. Всего известно более 100 типов, но их все делят на 2 группы:
1) Физиологические гемоглобины;
2) Патологические (аномальные).
К физиологическим гемоглобинам относятся:
- Hb P – примитивный гемоглобин, имеет место быть у 1-2 недельного эмбриона;
- Hb F – фетальный или гемоглобин плода, к моменту рождения составляет около 70% всего гемоглобина в крови;
- Hb A, Hb A2, Hb A3 – это гемоглобины взрослого организма. На Hb A приходится около 90-96%.
Физиологические типы гемоглобинов отличаются глобулиновой частью (АК-составом). Например Hb A содержит 2a и 2b субъединицы, а Hb F – 2a и 2g субъединицы.
К аномальным (возникающих при наследственных заболеваниях) гемоглобинам относятся:
- HbS – гемоглобин, сопутствующий серповидно-клеточной анемии. Отличается от нормального тем, что с N-конца в 6 положении b–цепи глутамин заменен на валин.
Миоглобин по сравнению с гемоглобином имеет третичную структуру, одну полипептидную цепь, один гем и может связывать одну молекулу кислорода. Гемоглобин и миоглобин функционируют вместе. Гемоглобин доставляет кислород из легких к тканям, а миоглобин перераспределяет его внутри клетки (доставляет к митохондриям).
Оба белка – гемопротеины, т.е. гемсодержащие белки.
Липид-белковые комплексы
Липид-белковые комплексы – сложные белки, простетическую часть которых составляют различные липидные компоненты. К таким компонентам относятся:
1. предельные и непредельные ВЖК. К предельным относятся стеариновая С17Н35СООН и пальмитиновая С15Н31СООН; к непредельным – олеиновая С17Н33СООН, линолевая С17Н31СООН, линоленовая С17Н29СООН и др.;
2. простые и смешанные триацилглицериды. К простым относится трипальмитин, а к смешанным – олеодистеарин :
СН2 - О - СО - С15Н31 СН2 - О - СО - С17Н33
| |
СН - О - СО - С15Н31 СН - О - СО - С17Н35
| |
СН2 - О - СО - С15Н31 СН2 - О - СО - С17Н35
3. фосфолипиды – сложные эфиры глицерина, ВЖК, остатка фосфорной кислоты и гидроксилсодержащего компонента (холин, серин, этаноламин, инозит и др.).
СН2 - О - СО - О - РО2Н - О - С2Н4 - NH2
|
СН - О - СО - С17Н35
|
СН2 - О - СО - С17Н35 [фосфатидилэтаноламин]
4. холестерин (холестерол), его эфиры. В основе холестерина лежит циклопентанпергидрофенантрен.

Связывание белковой и простетической части может осуществляется за счет:
- ионных связей, что характерно для связи фосфолипидов с белковой частью;
- гидрофобных взаимодействий между гидрофобным участком белковой молекулы и более гидрофобным участком простетической части.
Липопротеины. Делятся на:
- свободные липопротеины (ЛП) – хорошо растворимые в воде вещества, транспортные формы (доминируют белки);
- структурные протеолипиды – хорошо растворимые в органических растворителях.
К свободным ЛП относят транспортные ЛП крови. В составе этих частиц могут быть ВЖК, триацилглицерины, фосфолипиды, холестерин, холестериды и др. Наружная часть - p-полярные соединения (гидрофильные): белки, фосфолипиды, холестерин. В центре частицы локализованы холестериды, эфиры ВЖК, триацилглицириды, фосфолипиды [рис. этого]. Все их можно разделить по плотности и по электрофоретической подвижности:
1. хиломикроны (ХМ) – самые крупные и наименее плотные (r<1»0,95 кг/л) частицы. Состоят из 2% белка и 98% липидных компонентов (триглицериды). Образуются в кишечнике. Их количество резко возрастает после приема богатой липидами пищи;
2. ЛП очень низкой плотности (ЛПОНП) – по электрофоретической подвижности эта фракция предшествует b-ЛП (пре-b-ЛП). Их плотность =0,94 – 1,006 кг/л. Содержат повышенное количество белка. Образуются в печени, осуществляют транспорт триглицеридов из нее;
3. ЛП низкой плотности (ЛПНП) – по электрофоретической подвижности b-ЛП. Плотность от 1,006 до 1,06 кг/л;
4. ЛП высокой плотности (ЛПВП) – по электрофоретической подвижности a-ЛП. Плотность от 1,06 до 1,2 кг/л. Содержат примерно 60% белка.
По составу липидов, ЛП низкой и высокой плотности богаты холестерином и фосфолипидами.
Биологическая роль свободных ЛП сводится к транспортным функциям. За счет их гидрофильной оболочки происходит перенос различных веществ в клетки. Также они играют важную роль в диагностике патогенеза.
ЛПНП и ЛПОНП относятся к атерогенным ЛП, т.к. они богаты холестерином и являются крупными частицами, поэтому они застревают в сосудах и вызывают разрастание соединительной ткани.
ЛПВП – это антиатерогены, т.к. транспорт холестерина ими происходит из клеток в ткани.
Структурные липопротеины. Они плохо растворяются в воде, но хорошо в других растворителях. Участвуют в построении биомембран. Содержат 65-85% белка, находящийся в центре частицы и окружен липидами - поэтому их называют протеолипиды. В составе структурных ЛП важную роль играют фосфолипиды. Биологическая роль структурных ЛП заключается в транспорте веществ через клеточные мембраны, передаче нервного импульса и выполнении ими ферментативной функции.
НуклеопротеиныНуклеопротеины – это сложные белки, содержащие в качестве небольшой части нуклеиновые кислоты (до 65%).
НП состоят из 2-х частей: белковой (содержит гистоны и протамины, которые являясь основными белками, придают основные свойства) и простетической, представленной НК, сообщающими кислотные свойства. Взаимодействие между этими частями по ион-ионному механизму.
Все НП по составу НК можно разделить на 2 группы: рибонуклеопротеины (РНП) и дезоксирибонуклеопротеины (ДНП).
Состав НК:
НК – высокомолекулярные органические вещества, полинуклеотиды. Мономерами являются мононуклеотиды. Каждый мононуклеотид состоит из: углевода, азотистого основания и фосфорной кислоты. Так, РНК содержит b-D-рибофуранозу (рибозу), одно из 4-х возможных азотистых оснований (А, Г, Ц или У) и остаток фосфорной кислоты. ДНК содержит b-D-дезоксирибофуранозу (дезоксирибозу), одно из 4-х возможных азотистых оснований (А, Г, Ц или Т) и остаток фосфорной кислоты.


Строение азотистых оснований:
К группе пуриновых относятся аденин (6-аминопурин) и гуанин (2-амино-6-оксипурин). К группе пиримидиновых – урацил (2,4-диоксипиримидин), тимин (5-метилурацил) и цитозин (2-окси-4-аминопиримидин).




Схема образования нуклеотидов: [рис. схемы: аденин присоединяет рибозу и фосфорную к-ту, при этом выделяются 2 молекулы воды и образуется АМФ]. В клетке имеются нуклеотидфосфаты, дезоксинуклеотидфосфаты, трифосфаты (АТФ).
Структура нуклеиновых кислот:
Имеют несколько уровней структурной организации.
1. первичная структура. РНК и ДНК построены однотипно – представлены полинуклеотидной цепью, состоящей из отдельных мононуклеотодов, соединённых между собой 3’→5’-фосфодиэфирными связями. Эта связь образуется между фосфорным остатком одного мононуклеотида и 3’-ОН-группой пентозного остатка другого мононуклеотида. [рис. образования такой связи] Разные НК отличаются числом, порядком чередования и составом НК.
2. вторичная структура. По рентгеноструктурному анализу ДНК в 1953г Уотсон и Крик предложили модель строения ДНК, которая объясняла самовоспроизведение организмов, наследственную изменчивость. Вторичная структура представляет собой двойную спираль, состоящую из 2 полинуклеотидных цепей, закрученных вокруг одной общей оси. Эти цепи антипараллельны, т.е. одна идет в направлении 5’→3’, а другая 3’→5’. Пуриновому основанию одной цепи соответствует пиримидиновое основание другой цепи – эти основания комплиментарны друг другу, т.е. дополняют одно другое до целого. Между А и Т две водородные связи (А=Т), а между Г и Ц – 3 (ГºЦ).
Молекула спирализована на всем протяжении, гидрофобные участки внутри спирали, их плоскости перпендикулярны основаниям и параллельны друг другу. В вертикальном направлении возникают гидрофобные взаимодействия. Вторичная структура стабилизируется водородными связями и гидрофобными взаимодействиями.
Вторичная структура РНК более простая, представляет собой одну полинуклеотидную цепь, в которой спирализованы лишь некоторые участки. Вторичная структура РНК представлена в виде клеверного листа. Для тРНК известна третичная структура в форме буквы Г. [рис. РНК в виде клеверного листа]
Биологическая роль НК:
ДНК – основная часть её локализуется в ядре в виде ДНП в составе хроматина или хромосом делящихся клеток. Главная роль – хранение генетической информации, участие в процессе транскрипции в качестве матрицы для построения молекулы РНК.
Все РНК по функции делятся на:
- рРНК (рибосомальные), составляют до 80% в составе рибосом. Играют роль каркаса для объединения рибосом белков;
- мРНК (иРНК) – образуется в ядре (ядрышке). Переносит информацию из ядра в цитоплазму, является матрицей в процессе трансляции белка. последний кодон иРНК соответствует последней АК в белке;
- тРНК по своей форме напоминает форму клеверного листа и представляет собой полинуклеотидную цепь, которая составляет 3 петли и отдельные участки могут быть спирализованы. тРНК активирует аминокислоты и транспортирует их к месту биосинтеза белков, также участвует в трансляции. Имеет антикодоновый триплет – место, с помощью которого тРНК связывается с комплиментарным кодоном мРНК.
Углевод-белковые комплексыВ качестве простетической группы выступают углеводы. Все углевод-белковые комплексы делятся на гликопротеины и протеогликаны.
Гликопротеины (ГП)– комплекс белков с углеводными комплексами, где на долю углеводов приходится 2-20%. Углеводы представлены гетерополисахаридами, не имеющих системного строения (не происходит повторения углеводного фрагмента). До 15 видов моносахаридов могут присутствовать в этих гетерополисахаридах: глюкоза, галактоза, глюкозамин, галактозамин, глюкуроновая кислота, нейраминовая кислота (ПВК+маннозамин).






В качестве исключения к группе ГП относят ГП крови, которые определяют группу крови. В них на углеводную часть приходится до 80%, т.к. углеводный компонент не имеет регулярного строения. В этих белках между белковой и небелковой частями ковалентная связь, которая может быть разной по природе:
- гликозил-амидная: моносахариды связаны с амидной группой аспарагина (АСП). Такая связь в белках – иммуноглобулинах, гормонах и т.д.
- О-гликозидная: моносахарид образует гликозидную связь с гидроксилсодержащей АК – СЕР, ТРЕ. Такая связь характерна для муцина слюны, групповых веществ крови. Также этот тип связи может образовываться при взаимодействии моносахарида с ОН-группой окси-ПРО – в коллагеновых белках.
Свойства ГП:
1. термостабильность, т.е. при увеличении и уменьшении температуры белки не денатурируют;
2. плохо перевариваются трипсином и пепсином в ЖКТ;
3. специфичность молекулы – из них построены «узнающие участки» (узнают макромолекулы или участки поверхности клеток);
4. быстро выводятся из клеток и находятся в межклеточной жидкости или на поверхности клеточных мембран.
Биологическая роль ГП:
1. транспортная роль - связываются с различными веществами и переносят их. Пр.: ГП трансферин переносит ионы железа, церулоплазмин – переносит ионы меди, транскортин - гормоны.
2. каталитическая. ГП-ми являются многие ферменты. Пр.: пероксидаза, рибонуклеаза, энтерокиназа, холинэстераза.
3. защитная - входят в состав различных слизей, например муцин слюны.
4. свёртывающая. Пр.: фибриноген, протромбин участвуют в системе свертывания крови.
5. определяют группу крови. От строения нуклеотидных цепей ГП зависит специфичность ГП, определяющего группу крови.
6. ГП клеточной мембраны выделяются рецепторами и участвуют в механизмах межклеточного взаимодействия и узнавания.
7. регуляторная – некоторые гормоны являются ГП. Пр.: гонадотропный гормон, фолликулостимулирующий гормон.
Протеогликаны (ПГ). Углеводный компонент приходится 95%. Углеводные компоненты представлены линейными гетерополисахаридами системного строения. Их строение однотипно. Структурная единица – димер, состоящий из моносахаридов 2-х видов, которые, соединяясь, образуют структурную единицу полисахаридов. [рис. МС 1 и МС 2 соеденены между собой b-1,3-гликозидной связью, а сами эти фрагменты - собой b-1,4-гликозидной связью] МС1 – глюкуроновая кислота, МС2 – N-ацетилгексозамин.
Среди гетероплисахаридов, входящих в состав ПГ, основными являются: гиалуроновая кислота, хондроитинсульфаты, гепарин и др.
Гиалуроновая кислота. Полимер, структурная единица – димер, состоящий из глюкуроновой кислоты и N-ацетилглюкозамина.


Гепарин. Полимер, структурной единицей является димер, состоящий из глюкуроновой кислоты, связанной с N-ацетилглюкозамином, сульфатированым 4 или по 6 положению серной кислотой.

Гепарин не является структурным компонентом межклеточного матрикса. Он вырабатывается тучными клетками соединительной ткани и после их цитолиза выделяется в межклеточное пространство и кровеносное русло. В крови может соединяться с неспецифическими белками. Гепарин – естественный природный коагулянт, препятствует свёртыванию крови.
ПГ выполняют следующую роль:
1. роль «рессор» – смягчают нагрузки на суставные поверхности (в хрящах, суставных поверхностях);
2. ограничивают диффузию и проницаемость межклеточного вещества для различных молекул и крупных частиц. Гиалуроновая кислота подвергается гидролизу под действием фермента – гиалуронидазы. В норме, в здоровой ткани, фермент и субстрат находятся в равновесии. при патологических процессах количество фермента может увеличится, что приводит к гидролизу гиалуроновой кислоты и нарушению проницаемости. При этом разрываются b-1,4-гликозидные связи;
3. являются поливалентными анионами. Способны связывать большие количества ионов Na+ и др. катионов и участвуют в регуляции водно-солевого обмена;
4. Гиалуроновая кислота может присутствовать в свободном виде, например в хрусталике глаза, суставной жидкости. Она имеет консистенцию геля и выполняет различные функции.
ФосфопротеиныБелки, где в качестве простетической группы – фосфорная кислота. Присоединение фосфорной кислоты к полипептидной цепи идет с образованием сложноэфирной связи с АК СЕР или ТРЕ.


Типичными фосфопротеинами являются: казеин молока (1% фосфорной кислоты), желток куриного яйца (вителлин), икра рыб (ихтулин). Большое количество ФП содержится в нервных клетках. Для ФП характерен широкий диапазон функций в метаболизме. За счет фосфорилирования и дефосфорилирования ферментов происходит регуляция их активности.
ФЕРМЕНТЫ
Ферменты – это биологические катализаторы белковой природы. В одной клетке до 10 тыс. молекул фермента, которые катализируют 2000 ферментативных реакций. 1800 тыс. ферментов выделены, но их строение не расшифровано. Старое название ферментов – энзимы, а наука, их изучающая – энзимология.
По своей химической природе ферменты – это белки, они имеют несколько уровней структурной организации и обладают всеми другими свойствами белков. Очень большое количество ферментов не имеют 4-ую структуру, т.е. являются олигомерами.
Ферменты могут быть простыми и сложными. Простые состоят только из полипептидной цепи, а сложные имеют пептидную (апофермент) и небелковую части (кофермент). Далее идёт рис. [апофермент+кофермент=холофермент – обладает полноценной функциональной активностью]. По отдельности ни апофермент, ни кофермент, не выполняют функции так, как холофермент.
Строение ферментов. В пространственной структуре фермента условно выделяют ряд участков, которые выполняют соответствующие им функции. Активный центр (АЦ) – участок в молекуле фермента, где происходит связывание и химическое превращение субстрата (S). Субстрат – вещество, подвергающееся химическому превращению (например, для фермента лактатдегидрогеназы (ЛДГ) субстратом будет молочная кислота). В активном центре выделяется контактный участок и каталитический участок. Контактный участок – это место активного центра, в котором происходит связывание фермента с субстратом по принципу комплементарности, т.е. именно контактный участок обеспечивает специфическое сродство субстрата ферменту. Образовавшийся комплекс носит название фермент-субстратный комплекс. Каталитический участок (центр) – это место в активном центре фермента, где происходит химическое превращение субстрата [рис. изображён фермент-субстратный комплекс, а именно контактный участок, каталитический участок, активный центр и субстрат].
Если фермент – сложный белок, то обычно простетическая часть находится тоже в активном центре и участвует в формировании активного центра. Активный центр занимает небольшую часть молекулы фермента, обычно располагается в углублении, и в его образовании участвует небольшое число аминокислотных остатков (до 20). Аминокислотные остатки могут быть удалены друг от друга, но при формировании пространственной структуры фермента они располагаются в области активного центра.
В формировании активного центра могут участвовать остатки, несущие следующие функциональные группы: NH2 (ЛИЗ, АРГ), COOH (ГЛУ, АСП), OH (СЕР, ТРЕ), SH (ЦИС), имидазольное кольцо (ГИС). В качестве единиц, участвующих в формировании активного центра, могут выступать кофакторы - ионы металлов (Cu2+, Fe2+ и т.д.), а также коферменты. В сложном ферменте АК-остатки активного центра создают условия для правильной его конформации и помогают кофакторам в связывании, ориентации, а, следовательно, и в превращении субстрата.
Боковые группы остальных аминокислот не участвуют в образовании активного центра, но обеспечивают правильную пространственную конформацию активного центра и влияют на его реакционную способность.
Ряд ферментов могут содержать аллостерический центр. [рис. фермента с аллостерическим и активным центрами] Эти ферменты относят к аллостерическим ферментам. К аллостерическому центру присоединяются различные вещества, отличные по строению от субстрата. Эти вещества могут изменять конформацию активного центра, т.е. влиять на связывание и превращение субстрата, они называются аллостерическими эффекторами. Все аллостерические эффекторы делятся на положительные – активаторы, и отрицательные – ингибиторы.
Строение коферментовКоферменты в каталитических реакциях осуществляют транспорт различных групп атомов, электронов или протонов. Коферменты связываются с ферментами:
- ковалентными связями;
- ионными связями;
- гидрофобными взаимодействиями и т.д.
Один кофермент может быть коферментом для нескольких ферментов. Многие коферменты являются полифункциональными (например, НАД, ПФ). В зависимости от апофермента зависит специфичность холофермента.
Все коферменты делят на две большие группы: витаминные и невитаминные.
Коферменты витаминной природы – производные витаминов или химические модификации витаминов.
1 группа: тиаминовые – производные витамина В1. Сюда относят:
- тиаминмонофосфат (ТМФ);
- тиаминдифосфат (ТДФ) или тиаминпирофосфат (ТПФ) или кокарбоксилаза;
- тиаминтрифосфат (ТТФ).


ТПФ имеет наибольшее биологическое значение. Входит в состав декарбоксилазы кетокислот: ПВК, a-кетоглутаровая кислота. Этот фермент катализирует отщепление СО2.
Кокарбоксилаза участвует в транскетолазной реакции из пентозофосфатного цикла.
2 группа: флавиновые коферменты, производные витамина В2. Сюда относят:
- флавинмононуклеотид (ФМН);
- флавинадениндинуклеотид (ФАД).
Ребитол и изоалоксазин образуют витамин В2. Витамин В2 и остаток фосфорной к-ты образуют ФМН. ФМН в соединении с АМФ образуют ФАД.
[рис. изоалоксазиновое кольцо соединено с ребитолом, ребитол с фосфорной к-той, а фосфорная к-та – с АМФ]

ФАД и ФМН являются коферментами дегидрогеназ. Эти ферменты катализируют отщепление от субстрата водорода, т.е. участвуют в реакциях окисления–восстановления. Например СДГ – сукцинатдегидрогеназа – катализирует превращение янтарной к-ты в фумаровую. Это ФАД-зависимый фермент. [рис. COOH-CH2-CH2-COOH® (над стрелкой – СДГ, под – ФАД и ФАДН2) COOH-CH=CH-COOH]. Флавиновые ферменты (флавинзависимые ДГ) содержат ФАД, который в них является первоисточником протонов и электронов. В процессе хим. реакций ФАД превращается в ФАДН2. Рабочей частью ФАД является 2 кольцо изоалоксазина; в процессе хим. реакции идет присоединение двух атомов водорода к азотам и перегруппировка двойных связей в кольцах.

3 группа: пантотеновые коферменты, производные витамина В3 – пантотеновой кислоты. Входят в состав кофермента А, НS-КоА. Этот кофермент А является коферментом ацилтрансфераз, вместе с которой переносит различные группировки с одной молекулы на другую.
4 группа: никотинамидные, производные витамина РР - никотинамида:

Представители:
- никотинамидадениндинуклеотид (НАД);
- никотинамидадениндинуклеотидфосфат (НАДФ).
Коферменты НАД и НАДФ являются коферментами дегидрогеназ (НАДФ-зависимых ферментов), например малатДГ, изоцитратДГ, лактатДГ. Участвуют в процессах дегидрирования и в окислительно-восстановительных реакциях. При этом НАД присоединяет два протона и два электрона, и образуется НАДН2.

Рис. рабочей группы НАД и НАДФ: рисунок витамина РР, к которому присоединяется один атом Н и в результате происходит перегруппировка двойных связей. Рисуется новая конфигурация витамина РР + Н+]

5 группа: пиридоксиновые, производные витамина В6. [рис. пиридоксаля. Пиридоксаль+ фосфорная к-та= пиридоксальфосфат]

- пиридоксин;
- пиридоксаль;
- пиридоксамин.
Эти формы взаимопревращаются в процессе реакций. При взаимодействии пиридоксаля с фосфорной кислотой получается пиридоксальфосфат (ПФ).
ПФ является коферментом аминотрансфераз, осуществляет перенос аминогруппы от АК на кетокислоту – реакция переаминирования. Также производные витамина В6 входят как коферменты в состав декарбоксилаз АК.
Коферменты невитаминной природы – вещества, которые образуются в процессе метаболизма.
1) Нуклеотиды – УТФ, УДФ, ТТФ и т.д. УДФ-глюкоза вступает в синтез гликогена. УДФ-гиалуроновая к-та используется для обезвреживания различных веществ в трансверных реакциях (глюкоуронил трансфераза).
2) Производные порфирина (гем): каталаза, пероксидаза, цитохромы и т.д.
3) Пептиды. Глутатион – это трипептид (ГЛУ-ЦИС-ГЛИ), он участвует в о-в реакциях, является коферментом оксидоредуктаз (глутатионпероксидаза, глутатионредуктаза). 2GSH«(над стрелкой 2Н) G-S-S-G. GSH является восстановленной формой глутатиона, а G-S-S-G – окисленной.
4) Ионы металлов, например Zn2+ входит в состав фермента АлДГ (алкогольдегидрогеназы), Cu2+ - амилазы, Mg2+ - АТФ-азы (например, миозиновой АТФ-азы).
Могут участвовать в:
-присоединении субстратного комплекса фермента;
-в катализе;
-стабилизация оптимальной конформации активного центра фермента;
-стабилизация четвертичной структуры.
ИзоферментыИзоферменты – это изофункциональные белки. Они катализируют одну и ту же реакцию, но отличаются по некоторым функциональным свойствам в силу отличий по:
- аминокислотному составу;
- электрофоретической подвижности;
- молекулярной массе;
- кинетике ферментативных реакций;
- способу регуляции;
- стабильности и др.
Изоферменты – это молекулярные формы фермента, различия в аминокислотном составе обусловлены генетическими факторами.
Примеры изоферментов: глюкокиназа и гексокиназа.
 +АДФ
+АДФ
Гексокиназа может фосфорилировать любой шестичленный цикл, гексокиназа – только превращение глюкозы. После приёма пищи, богатой глюкозой, глюкокиназа начинает работать. Гексокиназа – стационарный фермент. Он катализирует реакцию расщепления глюкозы при низких её концентрациях, поступающих в организм. Отличаются по локализации (глюкокиназа – в печени, гексокиназа – в мышцах и печени), физиологическому значению, константе Михаэльса.
Если фермент – олигомерный белок, то изоформы могут получаться в результате различной комбинации протомеров. Например, лактатдегидрогеназа состоит из 4-х субъединиц. Н – субъединицы сердечного типа, М – мышечного. Может быть 5 комбинаций этих субъединиц, а, следовательно, и 5 изоферментов: НННН (ЛДГ1 – в сердечной мышце), НННМ (ЛДГ2), ННММ (ЛДГ3), НМММ (ЛДГ4), ММММ (ЛДГ5 – в печени и мышцах). [рис. эти 4 буквы в кружочки.
Надо отличать изоферменты от множественных форм ферментов. Множественные формы ферментов – это ферменты, которые модифицированы после своего синтеза, например фосфорилаза A и B.
Свойства ферментовОбщие черты ферментов и небиологических катализаторов:
1) и те, и другие катализируют только энергетически возможные реакции;
2) увеличивают скорость реакции;
3) не меняют направления реакции;
4) в ходе реакции не расходуются;
5) для обратимых процессов катализируют как прямую, так и обратную реакции, не смещая равновесия, а лишь ускоряя время его наступления.
Особые свойства ферментов:
1) высокая каталитическая активность. Металлы увеличивают скорость реакции в тысячи раз, а ферменты в миллионы раз. Например, уреаза ускоряет скорость реакции в 1014 раз. Каталаза ускоряет распад H2O2 в 1 млдр. раз! 2H2O2 ®2H2O +O2. Без катализатора выделения кислорода не видно. Металлический катализатор увеличивает скорость реакции в 1000 раз, а при добавлении каталазы – бурное вспенивание.
2) специфичность действия – наиболее характерная черта. Строение активного центра фермента, катализирующего реакции, различна. Структура активного центра фермента комплементарна структуре его субстрата, поэтому фермент из множества веществ присоединяет только свой субстрат – субстратная специфичность фермента.
Каждый фермент катализирует не любое превращение субстрата, а какое либо одно – специфичность пути превращения. Например, на АК ГИС действуют 2 фермента: гистидаза (отщепляет NH3) и гистидиндекарбоксилаза (отщепляет CO2).
Выделяют несколько видов специфичности:
а) абсолютная специфичность. Фермент действует только на один единственный субстрат. Пр.: уреаза разрушает мочевину: NH2-CO-NH2® (над стрелкой уреаза, под – вода) 2NH3+ CO2. Аргиназа катализирует распад аргинина.
б) групповая специфичность. Фермент действует на определённую связь в разных субстратах. Пр.: пептидазы разрывают пептидные связи [-NH-CH(R)-CO--NH-CH(R)-CO-]. Пепсин действует только на связи, образованные карбоксильной группой ароматических АК (ФЕН, ТИР, ТРИ). Эстеразы разрывают сложно-эфирную связь [-CO-NH-] в различных липидах. Гликозидазы действуют на гликозидную связь. Действие ферментов, обладающих групповой специфичностью, позволяет организму содержать небольшое количество ферментов.
в) стереоспецифичность. Фермент действует на определённый стереоизомер (D- и L-, цис- и транс-). Пр.: бутен-2-диовая кислота имеет 2 стереоизомера: транс-изомер или фумаровая к-та, и цис-изомер или малеиновая кислота.


Фумараза действует на фумаровую к-ту с превращением последней в яблочную.


В стереоспецифичности выделяют оптическую специфичность – избирательное действие ферментов на оптические изомеры. Например, под действием ЛДГ разрушается только L-форма молочной к-ты.


3) Влияние температуры (правило Вант-Гоффа). При увеличении температуры на 10 градусов скорость реакции увеличивается в 1,5-2 раза. Но для фермента это правило действует только до 40 градусов, т.к. дальше наступает тепловая денатурация фермента. Большинство ферментов в организме человека имеет оптимальную температуру 25-40 градусов [рис. графика: по оси х – температура, по у – процент активности. Рисуем горочку, оптимум – на 37-40°С].
Повышение активности фермента при увеличении температуры объясняется увеличением кинетической энергии реагирующих молекул, что приводит к увеличению числа столкновений между молекулами. При дальнейшем повышении температуры энергия становится чрезмерной, и внутри молекулы разрываются слабые связи – водородные, гидрофильные взаимодействия; происходит нарушение вторичной, третичной, четвертичной структуры фермента.
Ряд ферментов термостабильны, например, гликопротеины.
4) Влияние рН. Для поддержания третичной или четвертичной структуры фермента часто может быть необходимо наличие заряда на группе, удаленной от области связывания субстрата. Если же заряд этой группы меняется, то может происходить частичное развертывание белковой цепи, или компактизация, или диссоциация (олигомерные белки). Поэтому при отклонении рН от оптимального значения, фермент может потерять свою нативную структуру, в результате чего не происходит полноценного связывания активного центра с субстратом. Также при изменении рН может происходить изменение заряда на субстрате.
[рис. график. По х – рН, по у – процент активности. Рисуем горочку.]
Пепсин – 1.5-2, амилаза слюны - 6.8-7.2, трипсин - 7.5-8.6. Для большинства ферментов оптимум рН лежит в среде, близкой к нейтральной.
5) Скорость ферментативной реакции прямо пропорциональна кол-ву фермента (для небиологических катализаторов такой зависимости нет). Недостаток фермента в живом организме, например при неполноценном питании, генетических нарушениях, приводит к уменьшению скорости превращения веществ и наоборот.
6) Ферменты являются регулируемыми катализаторами. Так под действием различных веществ (активаторов и ингибиторов) меняется скорость ферментативной реакции.
Классификация и номенклатура ферментовСогласно международной классификации, принятой в 1961 году, все ферменты делятся на 6 классов по типу катализируемой реакции. Каждый класс делится на несколько подклассов. Классы:
1. оксидоредуктазы;
2. трансферазы;
3. гидралазы;
4. лиазы;
5. изомеразы;
6. лигазы (синтетазы).
1. Оксидредуктазы – ферменты, катализирующие окислительно-восстановительные реакции. Известно 480 этих ферментов, которые делятся на 17 подклассов.
а) аэробные дегидрогеназы (оксидазы). Отщепляют водород от субстрата и переносят его на молекулу кислорода. Пр.: НАДФН2+О2= Н2О2+НАДФ.
Если кислород непосредственно внедряется в субстрат, то ферменты – оксигеназы. Наибольшее значение имеют оксидазы аминокислот.
б) анаэробные дегидрогеназы – катализируют перенос водорода от субстрата на любой другой акцептор, кроме кислорода.
CH3CH2OH → СН3СН=О
[над стрелочкой алкоголь-ДГ; под стрелочкой НАД → НАДН2 ; этанол превращается в этаналь]
в) цитохромы – это ферменты, переносящие электроны.
г) пероксидазы – гемсодержащие оксидоредуктазы. Они отщепляют водород от субстрата и переносят его на Н2О2.
Пр.: каталазная реакция
2Н2О2 → 2Н2О+О2 [над стрелочкой - каталаза].
2. Трансферазы – ферменты, переносящие группы атомов от одного субстрата к другому. При этом один субстрат донор, а другой – акцептор. В зависимости от природы переносимых групп трансферазы делят на 8 подклассов:
- аминотрансферазы, переносят NH2;
- метилтрансферазы, переносят CH3;
- фосфотрансферазы, переносят PO3H2;
- ацилтрнсферазы.
Пр. переаминирование. [рис. Аланин + альфакетоглутаровая кислота под действием АлАТ и перидоксальфосфата получается ПВК и глутаминовая к-та].



3. Гидролазы – ферменты, катализирующие разрыв одинарных связей с участием воды, присоединяемой по месту разрыва связи. Т.е. они принимают участие в реакциях гидролиза. Все ферменты пищеварительного тракта относятся к гидролазам. Всего выделяют 460 гидролаз. В зависимости от типа разрываемых связей выделяют 11 подклассов:
- эстеразы – разрывают сложно-эфирную связь. Пр.: триацилглицерид+ 3H2O=(липаза) глицерин+ 3С17H35COOH [рис. этого. Стрелочкой показать сложноэфирные связи]
- пептидазы – разрывают пептидную связь. Пр.: АЛА-ГЛИ+ H2O = (дипептидаза) АЛА+ГЛИ [рис. этого. Стрелочкой показать пептидную связь]
- гликозидазы – разрывают гликозидные связи. Пр.: мальтоза+ H2O=(мальтаза) 2глюкозы [рис. этого. Стрелочкой показать гликозидную связь]
4) Лиазы. Эти ферменты осуществляют разрыв углеродных связей без участия воды. Выделяют:
- декарбоксилазы – катализируют отщепление CO2. Пр.:


- альдолазы – катализируют расщепление связи между атомами углерода. Пр.: фруктозо-1,6-дифосфат (6 атомов С)= (альдолаза) фосфоглицериновый альдегид (3 атома С)+ дигидроксиацетонфосфат (3 атома С).


- гидратазы – разрыв двойной связи с присоединением воды по месту разрыва двойной связи.
- ферменты, отщепляющие воду – реакция дегидратации. При этом образуется двойная связь.
5) Изомеразы – ферменты, катализирующие реакции изомеризации и обеспечивающие внутримолекулярную перестройку.


6) Лигазы (синтетазы) – катализируют образование более сложных веществ из более простых. При этом требуется энергия из вне. Обязательно участие АТФ или других трифосфатов (УТФ, ЦТФ и т.д.). [Пр. Аспарагиновая к-та + аммиак + АТФ под действием аспарагинсинтетазы превращается в аспарагин + АМФ + неорганический фосфат +энергия].


1) Существует тривиальная номенклатура – названия случайные, без системы и основания, например трипсин, пепсин, химотрипсин.
2) Рабочая номенклатура – название фермента составляется из названия субстрата или продукта реакции, типа катализируемой реакции и окончание –аза, например лактатдегирогеназа.
3) Систематическая, научная - L-лактат-НАД-оксидредуктаза.
4) Все ферменты имеют цифровой шифр, например ЛДГ - 1.1.1.27.
Первая цифра говорит о типе катализируемой реакции, указывая на номер класса.
Вторая уточняет действие фермента – номер подкласса.
Третья указывает природу разрываемой связи в молекуле субстрата - подподкласс.
Четвёртая – порядковый номер фермента.
Современные представления о ферментативном катализе
Первая теория ферментативного катализа была выдвинута в начале 20 века Варбургом и Бейлисом. Эта теория предлагала считать, что фермент адсорбирует на себе субстрат, и называлась адсорбционной, но развития она не получила.
В основу современной теории положена теория Михаэлеса и Ментена. Ведущую роль в механизме ферментативного катализа играет образование фермент-субстратного комплекса. По этой теории весь процесс катализа можно разделить на 3 этапа:
[рис. E+S«ES®ES*®ES**®EP®E+P]
1 этап: образование фермент-субстратного комплекса (на рис. до ES включительно). Происходит диффузия субстрата к ферменту и субстрат, в соответствии с принципом комплиментарности, связывается с активным центром фермента – образуется фермент-субстратный комплекс. Реагенты связаны слабыми связями, т.е. водородными, ионными, гидрофобными, в некоторых случаях и ковалентными. Эта стадия непродолжительна, зависит от концентрации субстрата и от скорости диффузии его к активному центру. Энергия активации исходных веществ при этом изменяется незначительно. На этой стадии проявляется эффект концентрирования субстрата на поверхности фермента – эффект ориентации.
2 этап: (на рис. от ES до EP включительно) происходит последовательное преобразование первичного фермент-субстратного комплекса в 1 или несколько активированных. Эта стадия наиболее медленна, ее длительность зависит от величины энергии активации данной реакции. В эту стадию происходит разрыв старых связей и образование новых, при этом энергия активации значительно снижается. По продолжительности эта стадия является лимитирующей для всего процесса.
На этой стадии проявляется эффект вынужденного соответствия – эффект «дыбы»: субстрат под действием фермента претерпевает изменения, делающие его более доступным для воздействия каталитического участка активного центра фермента. Одновременно с этим происходит изменение конформации фермента в большей степени в активном центре.
3 этап: отделение продуктов от активного центра фермента и диффузия их в окружающую среду. Эта стадия непродолжительна, ее скорость определяется скоростью диффузии продуктов в окружающую среду.
[график: по оси х – ход реакции, по у – энергия активации; рисуем горизонтальную прямую от середины оси у, затем горочку и окускаемся ниже прямой. пририсовываем меньшую горочку пунктиром]
Молекулярные эффекты действия ферментов1) Эффект концентрирования – это адсорбирование на поверхности молекулы фермента молекул реагирующих веществ, т.е. субстрата, что приводит к их лучшему взаимодействию. Пр.: электростатическое притяжение – скорость реакции может возрасти в 103 раз.
2) Эффект ориентации – это специфическое связывание субстрата с контактными участками активного центра фермента, которое обеспечивает взаимную ориентацию молекул субстрата и их сближение для более выгодного воздействия каталитических групп в активном центре. За счет эффекта ориентации скорость реакции возрастает в 103-104 раз. [рис. эффекта ориентации: поворот двух кругов вырезами друг к другу]
3) Эффект натяжения (теория «дыбы»). Субстрат до присоединения к ферменту находится в расслабленной конформации, а после связывания с ферментом деформируется или растягивается. Места деформации легче атакуются каталитическим центром фермента. [рис. эффекта дыбы: субстрат растягивается над ферментом]
4) Эффект вынужденного соответствия (прилегания). Не только субстрат претерпевает изменение конформации, но и фермент, особенно в активном центре, после связывания субстрата меняет свою конформацию, которая становится более комплементарной субстрату.
Теория Фишера: фермент подходит к субстрату как ключ к замку.
Теория Котланда: фермент и субстрат взаимодействуют между собой по принципу рука–перчатка. Истинная комплементарность фермента к субстрату достигается после изменения конформации и субстрата и фермента.
Теория кислотно-основного катализаВ составе активного центра фермента имеются как кислые, так и основные функциональные группы. В результате этого фермент проявляет в ходе катализа кислотно-основные свойства, т.е. играет как роль донора, так и роль акцептора протонов. Кислотно-основной катализ характерен для гидролаз, лиаз, изомераз.
При закреплении субстрата в активном центре на его молекулу влияют электрофильные и нуклеофильные группы каталитического участка, что вызывает перераспределение электронной плотности в субстрате. Такое перераспределение облегчает перестройку и разрыв связей в молекуле субстрата.
Пр.: Реакция превращения ацетилхолина в холин. На первом этапе между СОО- глутамина и N+ ацетилхолина возникает ионная связь, и возникает фермент-субстратный комплекс. Начинается вторая стадия.
После образования фермент-субстратного комплекса в действие вступают остальные аминокислоты, остатки активного центра. Между углеродом С=О группы ацетилхолина и кислородом ОН-группы серина возникает взаимодействие, т.е. возникает водородная связь между кислородом ацетилхолина и ОН-группы тирозина – эффект «дыбы».
Затем гистидин оттягивает протоны от ОН-группы серина. Вследствие этого упрочняется сложноэфирная связь между серином и остатком уксусной кислоты. Одновременно происходит разрыв другой сложноэфирной связи в молекуле ацетилхолина и переход протона от тирозина к остатку холина.
На третьем этапе холин высвобождается из активного центра. Его место занимает вода. Эта вода располагается между карбонильным кислородом ацетильной группы и кислородом тирозина. Фермент освобождён от продуктов реакции и готов к следующему циклу. На первом и последнем этапе продолжительность этапа зависит от скорости диффузии субстрата к ферменту или от фермента соответственно. Вторая стадия очень часто является лимитирующей весь процесс. Именно на этой стадии происходит снижение энергии активации реагирующих веществ.
Также существует ковалентный катализ – когда субстрат ковалентно связывается в активном центре фермента перед его превращением.
Регуляция активности ферментовФерменты являются регулируемыми катализаторами. В качестве регуляторов могут выступать метаболиты, яды. Различают:
- активаторы – вещества, увеличивающие скорость реакции;
- ингибиторы – вещества, уменьшающие скорость реакции.
Активация ферментов. Различные активаторы могут связываться либо с активным центром фермента, либо вне его. К группе активаторов, влияющих на активный центр, относятся: ионы металла, коферменты, сами субстраты.
Активация с помощью металлов протекает по различным механизмам:
- металл входит в состав каталитического участка активного центра;
- металл с субстратом образуют комплекс;
- за счет металла образуется мости между субстратом и активным центром фермента.
Субстраты также являются активаторами. При увеличении концентрации субстрата скорость реакции повышается. по достижению концентрации насыщения субстрата эта скорость не изменяется.
Если активатор связывается вне активного центра фермента, то происходит ковалентная модификация фермента:
1) частичный протеолиз (ограниченный протеолиз). Таким образом активируются ферменты пищеварительного канала: пепсин, трипсин, химотрипсин. Трипсин имеет состояние профермента трипсиногена, состоящего из 229 АК остатков. Под действием фермента энтерокиназы и с добавлением воды он превращается в трипсин, при этом отщепляется гексапептид. Изменяется третичная структура белка, формируется активный центр фермента и он переходит в активную форму.
2) фосфорилирование - дефосфорилирование. Пр.: липаза+АТФ= (протеинкиназа) фосфорилированная липаза+АДФ. Это трансферная реакция, использующая фосфат АТФ. При этом осуществляется перенос группы атомов от одной молекулы к другой. Фосфорилированная липаза является активной формой фермента.
Таким же путем происходит активация фосфорилазы: фосфорилаза B+ 4АТФ= фосфорилаза А+ 4АДФ
Также при связывании активатора вне активного центра происходит диссоциация неактивного комплекса «белок-активный фермент». Например, протеинкиназа – фермент, осуществляющий фосфорилирование (цАМФ-зависимое). Протеинкиназа – это белок, имеющий четвертичную структуру и состоящий из 2-х регуляторный и 2-х каталитических субъединиц. R2C2+2цАМФ=R2цАМФ2+ 2С. Такой тип регуляции называется аллостерической регуляцией (активацией).
Ингибирование ферментов. Ингибитор – это вещество, вызывающее специфическое снижение активности фермента. Следует различать ингибирование и инактивацию. Инактивация – это, например, денатурация белка в результате действия денатурирующих агентов.
По прочности связывания ингибитора с ферментом ингибиторы делят на обратимые и необратимые.
Необратимые ингибиторы прочно связаны и разрушают функциональные группы молекулы фермента, которые необходимы для проявления его каталитической активности. Все процедуры по очистке белка не влияют на связь ингибитора и фермента. Пр.: действие фосфорорганических соединений на фермент – холинэстеразу. Хлорофос, зарин, зоман и др. фосфорорганические соединения связываются с активным центром холинэстеразы. В результате происходит фосфорилирование каталитических групп активного центра фермента. В следствии молекулы фермента, связанные с ингибитором, не могут связываться с субстратом и наступает тяжелое отравление.
Также выделяют обратимые игнибиторы, например прозерин для холинэстеразы. Обратимое ингибирование зависит от концентрации субстрата и ингибитора и снимается избытком субстрата.
По механизму действия выделяют:
- конкурентное ингибирование;
- неконкурентное ингибирование;
- субстратное ингибирование;
- аллостерическое.
1) Конкурентное (изостерическое) ингибирование – это торможение ферментативной реакции, вызванное связыванием ингибитора с активным центром фермента. При этом ингибитор имеет сходство с субстратом. В процессе происходит конкуренция за активный центр: образуются фермент-субстратные и ингибитор-ферментные комплексы. E+S®ES® EP® E+P; E+I® E. Пр.: сукцинатдегидрогеназная реакция [рис. COOH-CH2-CH2-COOH®(над стрелкой СДГ, под ФАД®ФАДН2) COOH-CH=CH-COOH]. Истинным субстратом этой реакции является сукцинат (янтарная к-та). Ингибиторы: малоновая к-та (COOH-CH2-COOH) и оксалоацетат (COOH-CO-CH2-COOH). [рис. фермента с 3 дырками+ субстрат+ ингибитор= комплекс ингибитора с ферментом]
Пр.: фермент холинэстераза катализирует превращение ацетилхолина в холин: (CH3)3-N-CH2-CH2-O-CO-CH3® (над стрелкой ХЭ, под – вода) CH3СOOH+(CH3)3-N-CH2-CH2-OH. Конкурентными ингибиторами являются прозерин, севин.
2) Неконкурентное ингибирование – торможение, связанное с влиянием ингибитора на каталитическое превращение, но не на связывание фермента с субстратом. В этом случае ингибитор может связываться и с активным центром (каталитический участок) и вне его.
Присоединение ингибитора вне активного центра приводит к изменению конформации (третичной структуры) белка, вследствие чего изменяется конформация активного центра. Это затрагивает каталитический участок и мешает взаимодействию субстрата с активным центром. При этом ингибитор не имеет сходства с субстратом и это ингибирование нельзя снять избытком субстрата. Возможно образование тройных комплексов фермент-ингибитор-субстрат. Скорость такой реакции не будет максимальной.
К неконкурентным ингибиторам относят:
- цианиды. Они связываются с атомом железа в цитохромоксидазе и в результате этого фермент теряет свою активность, а т.к. это фермент дыхательной цепи, то нарушается дыхание клеток и они гибнут.
- ионы тяжёлых металлов и их органические соединения (Hg, Pb и др.). Механизм их действия связан с соединением их с различными SH-группами. [рис. фермента с SH-группами, иона ртути, субстрата. Все это соединяется в тройной комплекс]
- ряд фармакологических средств, которые должны поражать ферменты злокачественных клеток. Сюда же относятся ингибиторы, использующиеся в сельском хозяйстве, бытовые отравляющие вещества.
3) Субстратное ингибирование – торможение ферментативной реакции, вызванное избытком субстрата. Происходит в результате образования фермент-субстратного комплекса, неспособного подвергаться каталитическому превращению. Его можно снять и уменьшить концентрацию субстрата. [рис. связывания фермента сразу с 2 субстратами]
4) Аллостерическое ингибирование – торможение ферментативной реакции, вызванное присоединением аллостерического ингибитора в аллостерическом центре аллостерического фермента. Такой тип ингибирования характерен для аллостерических ферментов, имеющих четвертичную структуру. В качестве ингибиторов могут выступать метаболиты, гормоны, ионы металлов, коферменты.
Механизм действия:
а) присоединение ингибитора к аллостерическому центру;
б) изменяется конформация фермента;
в) изменяется конформация активного центра;
г) нарушается комплиментарность активного центра фермента к субстрату;
д) уменьшается число молекул ES;
е) уменьшается скорость ферментативной реакции.
[рис. фермент с 2 дырками, к одной аллостерический ингибитор и вторая меняет форму]
К особенностям аллостерических ферментов относят ингибирование по отрицателтной обратной связи. A®(E1)B®(E2) C®(E3) D (от D стрелочка к стрелке между А и В). D – метаболит, действующий как аллостерический ингибитор на фермент Е1.
Обмен веществОбмен веществ (метаболизм) – это совокупность физиологических и биохимических процессов, обеспечивающих жизнедеятельность организма во взаимосвязях с внешней средой, направленных на самовоспроизведение и самосохранение.
К физиологическим процессам относятся пищеварение, всасывание, внешнее дыхание, выделение и др.; к биохимическим – химические превращения белков, жиров, углеводов, поступающих в организм в виде пищевых веществ. Особенностью биохимических процессов является то, что они осуществляются в ходе ряда ферментативных реакций. Именно ферменты обеспечивают определенную последовательность, места и скорость реакций.
По направленности все химические превращения делят на:
а) диссимиляция (катаболизм) – распад веществ до более простых с переходом энергии связей вещества в энергию макроэргических связей (АТФ, НАД·Н, др.);
б) ассимиляция (анаболизм) – синтез более сложных веществ из более простых с затратой энергии.
Биологическое значение этих двух процессов состоит в том, что при расщеплении веществ освобождается заключенная в них энергия, которая обеспечивает все функциональные возможности организма. В то же время, при распаде веществ образуются "строительные материалы" (моносахариды, АК, глицерин и др.), которые затем используются в синтезе специфических организму веществ (белков, жиров, углеводов и др.).
[СХЕМА] Над горизонтальной линией (во внешней среде) – "белки, жиры, углеводы", от них стрелка вниз под линию (внутри организма) к надписи "диссимиляция", от последней четыре стрелки: две вверх к надписям над линией "теплота" и "конечные продукты"; одна стрелка вправо к надписи "промежуточные вещества (метаболиты)", от них к "ассимиляция", затем к "собственные белки, жиры, углеводы"; одна стрелка вниз к надписи "энергия АТФ", от нее – к "мышечное сокращение, проведение нервного импульса, секреция и др." а также наверх к "теплота" и "ассимиляция".
Диссимиляция белков, жиров и углеводов протекает по-разному, но в разрушении этих веществ есть ряд общих этапов:
1) Этап переваривания. В ЖКТ белки распадаются до АК, жиры – до глицерина и ВЖК, углеводы – до моносахаридов. Нарабатывается большое количество неспецифических веществ из специфических, поступающих извне. За счет переваривания в ЖКТ выделяется около 1% химической энергии веществ. Этот этап необходим для того, чтобы вещества, поступившие с пищей, смогли всосаться.
2) Этап межуточного обмена (тканевой обмен веществ, метаболизм). На клеточном уровне он распределяется на анаболизм и катаболизм. Образуются и превращаются промежуточные вещества обмена веществ – метаболиты. При этом мономеры, образовавшиеся на этапе переваривания, распадаются с образованием небольшого (до пяти) ключевых промежуточных продуктов: ЩУК, альфа-КГ, ацетил-КоА, ПВК, альфа-глицерофосфат. Выделяется до 20% энергии веществ. Как правило, межуточный обмен происходит в цитоплазме клеток.
3) Окончательный распад веществ с участием кислорода до конечных продуктов (СО2, Н2О, азотсодержащие вещества). Выделяется около 80% энергии веществ.
Все рассмотренные этапы отражают лишь главные формы обменных процессов. Как на втором, так и на третьем этапах выделяющаяся энергия накапливается в виде энергии химических связей макроэргических соединений (это вещества, имеющие хотя бы одну макроэргическую связь, напр., АТФ, ЦТФ, ТТФ, ГТФ, УТФ, АДФ, ЦДФ, …, креатинфосфат, 1,3-дифосфоглицериновая кислота). Так, энергия связи последнего фосфата в молекуле АТФ составляет около 10-12 ккал/моль.
Биологическая роль обмена веществ:
1. аккумуляция энергии при распаде химических соединений;
2. использование энергии для синтеза собственных веществ организма;
3. распад обновляемых структурных компонентов клетки;
4. происходит синтез и распад биомолекул специального назначения.
Обмен белков Переваривание и всасывание белковФункции белков многообразны, но особенно выделяются структурная, каталитическая и энергетическая функции. Энергетическая ценность белка около 4,1 ккал/г.
Среди всех веществ, поступающих в организм, при недостатке какого-либо из них, например, углеводов, их могут в некоторой степени заменить белки и липиды, а при недостатке липидов – белки и углеводы. Белки же практически незаменимы – они содержат 10 незаменимых АК: ВАЛ, ЛЕЙ, ИЛЕ, ЛИЗ, МЕТ, ТРЕ, ТРИ, ФЕН, ГИС, АРГ. При недостатке незаменимых АК лимитируется (ограничивается) синтез белков организма. Причем, примерно 50% поступающих белков должны составлять белки растительного и 50% - животного происхождения; максимально подходит по аминокислотному составу белок куриного яйца, он содержит около 34% незаменимых АК.
Взрослый мужчина нуждается в ежедневном употреблении 80-100 г белка. В 100 г белка содержится около 16 г азота, поэтому 1 г азота – в 6,25 г белка.
Для характеристики белкового (азотного) обмена веществ используется понятие «азотистый баланс» – это разность между азотом пищи и азотом мочи. В норме азотистый баланс равен нулю; положителен в растущем организме, во время активной регенерации, беременности; отрицателен – в старческом возрасте, при патологии, голодании.
Превращение белков в органах пищеваренияВсе белки подвергаются действию гидролаз (третий класс ферментов), а именно пептидаз – они, как правило, вырабатываются в неактивной форме, а затем активируются путем частичного протеолиза.
► Ротовая полость. Гидролиза нет.
► Желудок. Действуют ферменты:
1) Пепсин (вырабатывается в виде пепсиногена, который активируется соляной кислотой и своей активной формой – пепсином). Пепсин имеет одну полипептидную цепь. Под действием HCl или в результате аутокатализа происходит частичный протеолиз пепсиногена: от его N-конца отщепляется 42-АК фрагмент. После этого формируется третичная структура с полноценным активным центром.
Роль соляной кислоты:
ü активирует пепсиноген (пепсин);
ü создает оптимальную рН для действия пепсина (рН=1,5-2,5);
ü бактерицидное действие (напр., при снижении кислотности желудочного сока – гниение, брожение, выделяются сероводород, органические кислоты);
ü денатурирует белки пищи, и они становятся более доступными для действия пептидаз;
ü способствует эвакуации желудочного содержимого.
Пепсин – эндопептидаза (т.е. разрывает внутренние пептидные связи), действующая на пептидные связи, образованные СООН-группой ароматических АК (ГИС, ТРИ, ТИР, ФЕН).
2) Гастриксин (рНоптим.=3,5) – эндопептидаза, рвет связи, образованные дикарбоновыми АК (ГЛУ, АСП).
В желудке образуются достаточно крупные пептиды – пептоны, или альбумозы.
► Кишечник. Действуют ферменты: трипсин, химотрипсин, эластаза, карбоксипептидаза, аминопептидаза, дипептидазы, энтеропептидаза. Процесс начинается с превращения трипсиногена в трипсин под действием энтеропептидазы. Затем трипсин катализирует активацию:
химотрипсиноген → химотрипсин,
прокарбоксипептидазы → карбоксипептидазы,
проэластаза → эластаза.
В результате действия указанных ферментов происходит полный гидролиз белков пищи.
1) химотрипсин вырабатывается в ПЖЖ (поджелудочной железе), активируется частичным протеолизом трипсином. Действует в кишечнике (рН=7,5-8,5) как эндопептидаза; рвет связи, образованные СООН-группой ароматических АК.
2) трипсин вырабатывается в ПЖЖ, активируется частичным протеолизом (отщепление 6 АК-фрагмента с N-конца. Действует в тонкой кишке как эндопептидаза, рвет связи, образованные СООН-группами диаминомонокарбоновыми кислотами (ЛИЗ, АРГ).
3) эластаза вырабатывается в ПЖЖ, активируется частичным протеолизом трипсином. Действует в кишечнике как эндопептидаза; рвет связи, образованные АК пролином (ПРО).
4) карбоксипептидазы А и В вырабатываются в ПЖЖ, активируются частичным протеолизом трипсином. Действуют в кишечнике как экзопептидазы; рвут связи, образованные СООН-группами: А- ароматических и алифатических АК, В- ЛИЗ и АРГ.
5) аминопептидаза вырабатывается слизистой кишечника, активируется ионами Zn2+ и Mn2+. Действует в кишечнике как экзопептидаза; отщепляет по одной АК с N-конца.
6) дипептидазы вырабатываются слизистой кишечника, активируются ионами Co2+ и Mn2+. Действуют в кишечнике как экзопептидазы; расщепляют дипептиды.
7) эндопептидаза вырабатывается в кишечнике, активирует трипсиноген.
В кишечнике происходит полный гидролиз белков.
Переваривание сложных белков и их катаболизм1. Гликопротеины гидролизуются с помощью гликозидаз (амилолитических ферментов).
2. Липопротеины – с помощью липолитических ферментов.
3. Гемсодержащие хромопротеины: порфириновое кольцо не расщепляется, а выводится из организма с калом.
4. Нуклеопротеины. В кишечнике полинуклеотиды гидролизуются с помощью специфических нуклеаз до мононуклеотидов, которые под действием нуклеотидаз разрушаются до нуклеозидов и Фн. Затем под действием нуклеозидаз они распадаются до азотистых оснований и пентоз (рибоза или дезоксирибоза). Пуриновые азотистые основания разрушаются до мочевой к-ты, а пиримидиновые – до бета-аланина, аммиака NH3, углекислоты CO2.
Транспорт АК через цитолемму является активным и требует градиента Na+. Такой градиент создается ферментом Na+,K+-АТФ-азой, расположенным в цитолемме эпителиоцитов. Существует более пяти переносчиков АК:
ü для нейтральных алифатических АК (ЛЕЙ, ИЛЕ),
ü для ароматических АК,
ü для основных АК,
ü для кислых АК,
ü для АК пролина (ПРО).
А также имеется гамма-глутаминовый цикл (шунт), содержащий гамма-глутамилтранспептидазу (ГТТП, иначе гамма-глутамилтрансфераза – ГТТФ). Он имеет большое значение в определении функции печени, характеризуя степень усвоения АК гепатоцитами.
Гниение белков и обезвреживание его продуктовГниение белков – это бактериальный распад белковых веществ и АК под действием микрофлоры кишечника. Идет в толстой кишке, однако может наблюдаться и в желудке – при снижении кислотности в нем.
Образуются такие продукты:
а) токсичные: сероводород H2S, углекислота CO2, аммиак NH3, метан CH4, меркаптаны (CH3SH и его гомологи), бензол C6H6, крезол, индол, скатол и др.
б) нетоксичные: спирты (в т.ч. этиловый), амины, жирные к-ты, кетокислоты, витамины (напр., витамин B6).
Основные процессы гниения:
1. Декарбоксилирование (–СО2) обычно характерно для диаминомонокрбоновых кислот.
Напр., орнитин COOH-CH(NH2)-CH2-CH2-CH2-NH2 превращается в путресцин (то же, но без СООН-группы). Или лизин (ЛИЗ) – в кадаверин (это вроде первой реакции, но в цепи на одну CH2-группу больше).
Путресцин, кадаверин входят в состав трупных ядов. Они всасываются и частично выводятся с мочой и обезвреживаются в печени диаминооксидазой.
2. Дезаминирование. При гниении главным образом протекает восстановительное дезаминирование. Напр., аланин (АЛА) NH2-CH(CH3)-COOH + 2Н → СН3-СН2-СООН (пропионовая к-та) +NH3.
3. Десульфирование (тоже восстановительное).
Напр., цистеин (ЦИС) NH2-CH(CH2SH)-COOH + 2Н → АЛА + H2S.
Напр., метионин (МЕТ) NH2-CH(CH2-СН2-S-CH3)-COOH + 2Н → NH2-CH(CH2-CH3)-COOH (альфа-аминомасляная к-та) + НS-CH3 (метил-меркаптан).
4. Разрушение боковой цепи АК (ТИР, ТРИ).
Напр., тирозин (ТИР) + 4Н → крезол (пара-метилфенол) + NH3 + CO2 + CH4; крезол + 2Н → фенол + СН4.
Напр., триптофан (ТРИ) + 4Н → скатол (метилиндол) + NH2 + CO2 + CH4. Скатол +2Н→ индол + СН4.
Т.о., в процессе гниения АК образуются различные токсичные вещества, которые должны быть обезврежены в печени. В обезвреживании участвуют две системы:
1) УДФГК – уридиндифосфоглюкуроновая к-та (активная форма глюкуроновой к-ты)

2) ФАФС – 3"-фосфоаденозин-5’-фосфосульфат (активная форма серной к-ты) [рис. этого соединения].
Механизм обезвреживания – конъюгация (связывание) токсина с активной формой серной или глюкуроновой к-ты. Продукты конъюгации – нетоксичные вещества, которые могут выделяться с мочой.
Напр., обезвреживание фенола под действием УДФ-глюкуронил-трансферазы: фенол + УДФГК → (ТФ) фенилглюкуронид (фенил присоединяется по первому положению) + R-OH.
Напр., обезвреживание индола: Индол окисляется кислородом по 7-му положению, получается индоксил (это типа 7-оксииндол). Индоксил взаимодействует с ФАФС под действием арилссульфо-трансферазы с образованием индоксилсерной к-ты, которая с ионами калия дает индикан (калиевая соль индоксилсульфата).


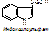
Определение индола и индикана в моче имеет диагностическое значение. Так, если отсутствует индол, то обезвреживающая функция печени в норме, а если при этом обнаруживается индикан, то в кишечнике активное гниение. Если же есть индол в моче, то имеется нарушение обезвреживающей функции печени.
Метаболизм аминокислотФонд АК организма пополняется за счет процессов:
1) гидролиза белков пищи,
2) гидролиза тканевых белков (под действием катепсинов лизосом).
Расходуется АК-фонд на процессы:
ü синтез заменимых АК,
ü синтез собственных белков,
ü синтез азотсодержащих веществ (урины, пиримидины, холин, креатин и т.д.),
ü синтез углеводов (глюконеогенез),
ü синтез липидов из кетогенных АК,
ü распад до NH3, NH2-CO-NH2, мочевой к-ты и др.
Условно метаболизм АК в тканях можно распределить на общие пути и индивидуальные пути обмена АК.
Общие пути обмена веществ1. Переаминирование (открыто в 1937 г. Браунштейном и Крицмом).


Роль: синтез заменимых АК, участие в непрямом дезаминировании АК. Определение АлАТ и АсАТ в крови имеет большое диагностическое значение. Так, через 5 часов после инфаркта миокарда АсАТ увеличивается в 20-30 раз, через 48 часов – АлАТ и АсАТ снижаются до нормы, еще через 24 часа повышается АлАТ. Также АлАТ повышается при патологии печени.
2. Дезаминирование (ДА) АК:
ü восстановительное ДА – под действием микрофлоры кишечника,
ü гидролитическое ДА – с участием воды,
ü внутримолекулярное ДА – с образованием непредельной к-ты,
ü окислительное ДА – характерно для тканей организма. Оно бывает прямым и непрямым.
Прямое ДА идет с участием дезаминаз (оксидаз). NH2-CHR-COOH → NH=CR-COOH (иминокислота), при этом ФМН→ФМН·Н2, который затем восстанавливает кислород до пероксида водорода; последний расщепляется каталазой. А иминокислота гидролизуется до альфа-кетокислоты и аммиака.
Непрямое ДА (или транс-ДА) идет в два этапа: 1) переаминирование (см. выше); 2) дезаминирование ГЛУ «α-КГ + NH3, над стрелочкой глутамат-ДГ, под стрелочкой – НАД→НАД·Н2.
3. Декарбоксилирование АК – процессы образования биогенных аминов, обладающих биологической активностью:
ГИС → (гистидил-ДК, ПФ) гистамин,


ТИР → (оксигеназа, +1/2О2) ДОФА (диоксифенилаланин) → (ДК, ПФ, -СО2)дофамин,






ТРИ → (оксигеназа, +1/2О2) 5-окситриптофан → (ДК, ПФ, -СО2) серотонин,
ГЛУ → гамма-аминомасляная к-та (ГАМК).


Дофамин и ГАМК – тормозные нейромедиаторы, гистамин – тканевой гормон. Серотонин является местным регулятором в функции периферических органов.
Образование конечных азотистых продуктовВ сутки распадается около 1-2% всех белков организма, что составляет в среднем 500 г. Из них 80% (400 г) идут на ресинтез организм-специфичных белков, а 20% (100 г) подвергаются непрямому дезаминированию с образованием конечных продуктов – кетокислот и аммиака (они содержат 10-16 г азота).
Временное обезвреживание аммиакаАммиак токсичен (50 мг аммиака убивает кролика, при этом [NH3]=0,4-0,7 мг/л). Поэтому в тканях аммиак обезвреживается временными путями:
1) в основном – образованием амидов дикарбоновых кислот. Напр., ГЛУ + NH3 → ГЛН (над стрелочкой "глутаминсинтетаза", под стрелочкой – АТФ → АДФ + Фн). Аналогично АСП → АСН.
→ 
 →
→
2) восстановительное аминирование кетокислот. Этот путь и дает токсичность аммиака (из-за уменьшения кол-ва кетокислот).
Такой азот (в виде конъюгатов аммиака) посупает в печень, где происходит окончательное обезвреживание аммиака – образование мочевины. Небольшое количество аминов отдают аммиак в почках, где он сразу синтезируется в мочу, где соединяется с протонами, образуя ионы аммония, которые выводятся с мочой. (В крови NH4+ нет!)
Орнитиновый цикл мочевинообразованияМочевина содержит 80-90% всего азота мочи. В сутки образуется 25-30 г мочевины NH2-CO-NH2.
1. NH3 + CO2 + 2АТФ + Н2О → H2N-CO-OPO3H2 + 2АДФ + Н3РО4 (над стрелочкой карбамоилфосфатсинтетаза). Образуется карамоилфосфат.
2. NH2-(CH2)3-CH(NH2)-COOH (это орнитин) + H2N-CO-OPO3H2 (это карбамоилфосфат) ® (орнитинкарбамоил-ТФ, – Н3РО4) NH2-CO-NH-(CH2)3-CH(NH2)-COOH (это цитруллин)
Далее: NH2-CO-NH-(CH2)3-CH(NH2)-COOH (это цитруллин) +COOH-CH(NH2)-CH2-COOH (это АСП)® аргининосукцинатсинтетаза, АТФ→АМФ+ФФн) СООН-СН2-СН(СООН)-NH-C(=NH)-NH-(CH2)3-CH(NH2)-COOH (это аргининосукцинат) ®(аргининосукцинатлиаза) СООН-СН=СН-СООН (это фумарат, который идете в ЦТК)) +NH2-C(=NH)-NH-(CH2)3-CH(NH2)-COOH (это аргинин).
Затем: NH2-C(=NH)-NH-(CH2)3-CH(NH2)-COOH (это аргинин)® (аргиназа, +Н2О) NH2-C(=O)-NH2 (это мочевина) + NH2-(CH2)3-CH(NH2)-COOH (это орнитин)
Т.о., для синтеза мочевины необходимо два азота – один из АСП и один из ГЛУ, ГЛН, АЛА; а также 3 АТФ и 1 СО2.
Синтез и распад нуклеотидовОсобенности обмена нуклеотидов:
1. Ни сами нуклеотиды, ни азотистые основания, поступающие с пищей, не включаются в синтез нуклеиновых кислот и нуклеотидов организма. Т.е., нуклеотиды пищи всегда распадаются до конечных продуктов.
2. Нуклеотиды синтезируются клетками организма заново, при этом не используются нуклеотиды пищи.
Биосинтез пуриновых оснований – из аминокислот: ГЛИ, АСП, ГЛН, два атома углерода – из производного фолиевой к-ты, один – из углекислоты.
Пиримидиновые основания также образуются из АК.
Распад нуклеотидов аналогичен в тканях и в ЖКТ:
нуклеопротеины ® нуклеиновые к-ты + АК (над стрелкой – протеазы); ДНК / РНК ®(ДНК-аза / РНК-аза, +вода) полинуклеотиды ®(полинуклеотидазы) мононуклеотиды ®(нуклеотидазы, -Фн) нуклеозиды
Распад нуклеозидов сходен в тканях и в ЖКТ, но имеются различия в окислении пуриновых и пиримидиновых нуклеозидов.
А. Окисление пуриновых нуклеозидовАденозин® (аденозиндезаминаза, +Н2О, –NH4+) инозин® (пуриннуклеозидфосфорилаза, +Фн –рибозил-1-Ф) гипоксантин (6-оксопурин) ® (ксантиноксидаза, +О2, +Н2О → Н2О2) ксантин (или 2,6-диоксопурин) ®(ксантиноксидаза, +О2, +Н2О → Н2О2) мочевая к-та (или 2,6,8-триоксопурин)
Мочевая к-та – конечный продукт распада пуриновых оснований.
Б. Распад пиримидиновых нуклеозидов
Они распадаются до бета-аланина, который окисляется до СО2 и NH3. А также до бета-амилоизобутирата. Аммиак, образующийся при распаде, обезвреживается с образованием мочевины.
Повышение содержания мочевой к-ты в крови (гиперурикемия) наблюдается при повышенном распаде пуриновых нуклеотидов (как поступающих с пищей, так, возможно, и собственных).
Однако, мочевая к-та плохо растворима в кислой среде, поэтому может кристаллизоваться и осаждаться в суставах (подагра), в мочевыводящих путях (мочекаменная болезнь). Лечение – растворение с помощью содового питья (образуются хорошо растворимые ураты).
Образование креатинина
Креатинин образуется из креатина – азотистого соединения мышечной ткани. Креатин синтезируется в печени из АК: АРГ, МЕТ, ГЛИ. Затем креатин поступает в мышцу, где связывает фосфат с образованием креатинфосфата.
Иногда креатинфосфат "теряет" фосфат, тогда образуется креатинин – конечный продукт распада креатина (в креатинине образуется связь между азотом аминогруппы и углеродом карбокси-группы).

Количество креатинина в моче определяется как показатель клубочковой фильтрации почек (он не реабсорбируется). Также определяют клиренс креатинина – сравнение содержания его в крови и в моче.
Все конечные продукты, содержащиеся в крови, составляют остаточный азот крови – это азот, остающийся в растворе после осаждения белков. В норме остаточный азот 14-28 ммоль/л. Он состоит из азота промежуточных продуктов (пептиды, АК, билирубин, нуклеотиды, креатин, индол) и азота конечных продуктов (мочевина, мочевая кислота, креатинин, индикан).
Образование конечных безазотистых продуктов
I. Превращение ПВК
В результате непрямого дезаминирования из АК образуются аммиак и кетокислоты.
ЛЕЙ ® кетокислоты ® жиры.
ИЛЕ, ЛИЗ, ФЕН, ТРИ, ТИР ® углеводы и жиры.
АЛА, ЦИС, СЕР, ГЛИ, ТРЕ ® ПВК.
ГЛУ ® альфа-КГ.
АСП ® оксалоацетат (ЩУК).
Пировиноградная к-та (ПВК) подвергается окислительному декарбоксилированию (Ох-ДК), протекающему в четыре реакции. При этом молекула ПВК [СН3-СО-СООН] превращается в ацетил-КоА [СH3-CO-S-KoA] и углекислоту [СО2]. Пируват-дегидрогеназный комплекс ферментов: пируватДК, ацетилТФ, ДГ липоевой к-ты. Коферменты: НАД, ФАД, ТДФ, HSKoA, липоевая к-та. НАД переходит в восстановленную форму НАД·Н2, который поступает в дыхательную цепь митохондрий, где дает 3 АТФ.
Ацетил-КоА вступает в ЦТК (цикл трикарбоновых кислот, или цикл Кребса, или цикл лимонной кислоты).
II. Цикл Кребса:

[НАД·Н2 поступает в дыхательную цепь митохондрий, где каждая молекула дает три АТФ; ФАД·Н2 – аналогично, но дает две АТФ]
Суммарно ЦТК выглядит так:
СН3-СО-SКoA + 2H2O + Фн + АДФ ®2СО2 + 3НАД·Н2 + ФАД·Н2 + АТФ + HSКоА
Функции ЦТК:1. катаболическая – распад белков, жиров, углеводов;
2. интегративная – взаимосвязь обменов Б, Ж, У;
3. анаболическая – использование метаболитов ЦТК в синтезе необходимых организму соединений (напр., альфа-КГ ® глу ; ЩУК ® асп);
4. энергетическая – выделение АТФ.
5. водород-генерирующая – образование восстановленных форм коферментов (3 НАД·Н2 + ФАД·Н2 ® 4 Н2)
III. Биологическое окисление.
Биологическое окисление – это совокупность реакций окисления, протекающих в живом организме.
Лавуазье сравнивал биологическое окисление с «медленным горением», но это ограниченная аналогия, так как биологическое окисление:
а) протекает при низкой температуре,
б) протекает без появления пламени,
в) протекает в присутствии воды.
Существует несколько теорий биологического окисления:
1. Теория “активации” кислорода (Бах)
Образуются пероксиды: а) О=О®–О–О–
б) –О–О– + S ® SOO [треугольник, в углах которого три указанные буквы; S = субстрат; над стрелочкой реакции надпись “оксигеназы”]
в) SOO + S’ ® SO + S’O [фермент пероксидаза]
Эта теория не объясняет окисление в животных тканях.
2. Теория активирования водорода (Палладин)
В клетках животных окисление идет благодаря дегидрированию:
А·Н2 + Ко® А + Ко·Н2 [фермент дегидрогеназа]
Ко·Н2 + ½О2 ® Ко + Н2О
3. Современные представления (Палладин и Бах)
Биологическое окисление – это процесс переноса электронов. Если акцептором электронов выступает молекулярный кислород, то его называют “тканевым дыханием”:
RH2 ® R + 2H+ + 2e--
2H+ + 2e-- + ½ O2 ® H2O + 210 кДж
Биологическое окисление – многоступенчатый полиферментативный процесс, заключающийся в многократной передаче протонов и электронов по цепи ферментов. При этом химическая энергия выделяется небольшими порциями (постепенно, без взрывов).
Дыхательная цепь (ДЦ) (или Цепь Переноса Электронов – ЦПЭ, или Электрон-Транспортная Цепь – ЭТЦ)ДЦ – это конвейер по переносу электронов и протонов от восстановленного субстрата к кислороду.
Компоненты ДЦ:
1. Пиридинзависимые ДГ (НАД-, НАДФ-зависимые)
Рабочая часть – витамин РР (никотинамид)

НАД + 2Н+ + 2е ↔ НАД·Н2
2. ФАД-зависимые ДГ (кофермент в ДЦ – ФМН, а акцептор электронов непосредственно от субстрата – ФАД. Рабочая часть – изоалоксазин.

[При восстановлении к атомам азота при двойных связях, отмеченных стрелками, присоединяется по атому водорода, а двойная связь перемещается на общую грань колец В и С.]
3. Убихинон (Ko Q). Обладает о/в-свойствами благодаря кето-енольной таутомерии.
4. Цитохромы. Относятся к гемопротеинам, содержат атомы железа, переход степени (2↔3) окисления которого и обеспечивает транспорт электронов (протоны ими не транспортируются !!! ).
В ДЦ цитохромы расположены в следующей последовательности: b - c1 - c - a - a3 .
Совокупность цитохромов b и c1 называют КоQH-дегидрогеназой, т.к. они отщепляют атом водорода от убихинона (KoQ).
Цитохромы а и а3 – цитохромоксидазой (т.к. способствуют переносу электронов на молекулярный кислород).
Функционирование ДЦСубстрат·Н2 → НАД → ФМН → КоQ → 2b → 2c1 → 2c → 2a → 2a3 → O2 .
[До KoQ включительно переносятся 2 протона и 2 электрона, а по цепи цитохромов – только 2 электрона]
Существует и укороченная ДЦ, в которой субстрат окисляется ФАД-зависимой ДГ, отдающей затем 2 протона и 2 электрона непосредственно на убихинон.
Необходимо отметить, что АТФ выделяется на этапах: НАД→ФМН (в укороченной ДЦ эта молекула АТФ не выделяется!), b→c1 , a→a3 .
Вообще, молекула АТФ синтезируется если разница потенциалов между соседними компонентами цепи превышает 0,2 В, т.е. может выделиться энергия не менее 50 кДж/моль.
Окислительное фосфорилирование(хемиосмотическая теория Митчелла, 1961)
1. Мембраны митохондрий непроницаемы для протонов.
2. В результате процессов окисления в митохондрии формируется протонный потенциал (электрохимический градиент протонов).
3. Диффузия протонов обратно на внутреннюю поверхность мембраны сопряжено с фосфорилированием, которое осуществляется АТФ-синтетазой.
Сам процесс выглядит так:
1. НАД·Н-ДГ отдает пару электронов на ФМН-ДГ. Это позволяет ФМН принять пару протонов из матрикса с образованием ФМН·Н2. Пара протонов, принадлежащих НАД, выталкивается на цитоплазматическую поверхность внутренней мембраны митохондрий.
2. ФМН·Н2-ДГ выталкивает пару протонов на цитоплазматическую поверхность мембраны, а пару электронов отдает на убихинон (KoQ), который при этом получает способность присоединить пару электронов из матрикса с образованием KoQ·H2.
3. KoQ·H2 выталкивает пару протонов в цитоплазму, а электроны перебрасываются на кислород в матриксе с образованием воды. В итоге при переносе пары электронов из матрикса в межмембранное пространство перекачивается 6 протонов, что и ведет к созданию разницы потенциалов и разницы рН между поверхностями внутренней мембраны.
4. Разница потенциалов и разница рН обеспечивают движение протонов через протонный канал в обратном направлении.
5. Движение протонов ведет к активации АТФ-синтетазы и синтезу АТФ из АДФ и фосфата.
Окислительное фосфорилирование – это процесс образования АТФ из АДФ и ФН за счет энергии транспорта электронов в дыхательной цепи
Возможно разобщение окислительного фосфорилирования, например, при повреждении внутренней мембраны митохондрий. При этом происходит свободное окисление без фосфорилирования (т.е. без синтеза АТФ), сопровождающееся пирогенным эффектом (локальным повышением температуры). Таким образом на митохондиальную мембрану действует, например, 2,4-динитрофенол.
Альтернативные варианты биологического окисления
К ним относят оксигеназный путь окисления. Он не относится к энергоснабжающим клетку процессам, а используется для деградации (разрушения) метаболитов. Ферменты оксигеназного пути катализируют включение кислорода в субстрат.
Различают ди- и монооксигеназный пути.
Диоксигеназный путь содержит ферменты, которые включают в молекулу субстрата оба атома кислорода. Этот вариант встречается очень редко.
S + O2 → S·O2
Монооксигеназный путь – в молекулу субстрата включается один из атомов кислорода, а другой восстанавливается до воды. Для этого необходим еще косубстрат (донор электронов).
A-H + O2 + ZH2 → A-OH + Z + H2O
Для примера рассмотрим систему микросомального окисления. Она содержит цитохромы Р-450 и b5. Эта система играет большую роль в обезвреживании многих токсинов и лекарств путем их гидроксилирования.
R-H → R-OH
При этом часто образуется пероксид водорода, который разрушается каталазой.
Матричный биосинтез
Генетический код
Понятие о генетическом коде было введено в середине 50-х гг. Гамовым. Генетически код - это информация об аминокислотах (АК), записанная в виде последовательности нуклеотидов. Посредством 4-х нуклеотидов кодируется 20 основных АК.
Свойства генетического кода:
1. генетический код триплетный, т.е. одна последовательность состоит из 3-х нуклеотидов. Имеется 43 (64) варианта, а необходимо 20, поэтому генетический код имеет квазидуплетность – смысловую нагрузку для большинства АК несут первые 2 нуклеотида, а третий нуклеотид для некоторых АК не важен вообще, а для других имеет значение пуриновый он или пиримидиновый. И только для ТРИ и МЕТ важен 3 нуклеотид;
2. однозначность - один код (триплет) несет информацию только об одной АК;
3. вырожденность - одной АК соответствует несколько кодонов. Это происходит вследствие того, что кодонов 64, а АК - 20. Так, СЕР соответствует 6 кодонов, ГЛИ и АЛА - по 4 кодона. Большей части АК соответствует по 2 кода, только ТРИ и МЕТ кодируются 1 кодоном;
Также имеются кодоны, которые не несут смысловой нагрузки - нонсенс (бессмысленные) кодоны – терминирующие кодоны;
4. неперекрещиваемость - считывание информации идет от одного триплета к другому триплету последовательно;
5. универсальность - для всего живого генетический код един.
Т.о., в виде генетического кода записана информация об одной АК, а последовательность нуклеотидов (в виде триплетов) несет информацию о последовательности АК в полипептидной цепи.
Отрезок ДНК, несущий информацию о последовательности АК в одной полипептидной цепи, называется геном.
Функции гена:
1. хранение информации об одной полипептидной цепи;
2. передача информации из поколения в поколение клеток;
3. передача информации с ДНК на РНК - транскрипция (синтез РНК);
4. передача информации с РНК на последовательность АК в последовательность полипептидной цепи - трансляция (декодирование информации РНК в последовательность АК) - биосинтез белка.
Репликация (самоудвоение, биосинтез) ДНК
В 1953 г. Уотсон и Крик открыли принцип комплементарности (взаимодополняемости). Так, А=Т, а ГºЦ.
Условия, необходимые для репликации:
1. строительный материал - дезоксинуклеозидтрифосфаты (дАТФ, дГТФ, дЦТФ ТТФ);
2. энергия, которая выделяется из вышеперечисленных трифосфатов: дАТФ ®дАМФ + ФФн +Q;
3. ионы Мg2+, играющие стабилизирующую роль;
4 матрица - расплетенная двойная спираль ДНК. Это расплетение называется репликативной вилкой [рис. расплетенной ДНК и образовавщейся репликативной вилки];
5. репликативный комплекс ферментов:
- ДНК-раскручивающие белки;
- ДНК-полимераза;
- ДНК-лигаза.
Основные этапы репликации:
1. образование репликативных вилок при участии ДНК-раскручивающих белков, вызывающих разрыв водородных связей между комплементарными основаниями [рис. репликативной вилки, на одном краю которой сплошная линия, а на другом – фрагментами – это фрагменты Оказаки. Помечены 5’ и 3‘ концы];
2. синтез новых нитей ДНК при участии ДНК-полимеразы, катализирующей образование фосфодиэфирной связи между новыми нуклеотидами. Присоединение нуклеотидов идет в соответствии с принципом комплементарности. Синтез идет 5’-конца к 3’-концу. На одной цепи синтез происходит непрерывно, а на другой - прерывается с образованием коротких фрагментов. В результате на одной цепи образуются короткие фрагменты - фрагменты Оказаки;
3. соединение коротких фрагментов с помощью ДНК-лигазы с образованием дочерних нитей.
В результате репликации на одной материнской нити синтезируются 2 комплементарных дочерних ДНК. Т.е. из одной молекулы ДНК образуются 2 копии ДНК.
Репликация протекает в ядре и частично в митохондриях в синтетическую фазу митотического цикла (S фаза). Значение репликации состоит в передаче информации от ДНК матери к дочерней ДНК.
Транскрипция (передача информации с ДНК на РНК) или биосинтез РНК
При транскрипции, в отличие от репликации, информации передается с небольшого участка ДНК. Элементарной единицей транскрипции является оперон (транскриптон)- участок ДНК, подвергающийся транскрипции.
В опероне выделяют информативные участки - экзоны и неинформативные участки - интроны. В начале оперона выделяют промотор (P) - это начальный участок оперона, к которому присоединяется РНК-полимераза. Рядом с промотором располагается оператор (О) - регуляторная зона, место присоединения генов-регуляторов. В конце оперона располагается терминатор (Т) - участок, содержащий стоп-сигнал. [рис. нескольких последовательных квадратиков: в начале помечаем Р, затем О, а следом экзоны и интроны (какие-то закрашиваем). В конце - Т]
Необходимые условия для трансляции:
1. расплетенный участок ДНК – одна нить ДНК;
2. строительный материал, представленный нуклеотидами - рибонуклеозидтрифосфаты (АТФ, ГТФ, ЦТФ, УТФ);
3. энергия, которая выделяется из вышеперечисленных трифосфатов: АТФ ®АМФ + ФФн +Q;
4. ферменты - ДНК-зависимая РНК-полимераза.
Этапы трансляции:
1. инициация (начало);
2. элонгация (продолжение);
3. терминация (окончание).
После транскрипции идет процессинг (созревание) РНК.
1. Инициация - заключается в присоединении ДНК-зависимой РНК-полимеразы к промотору, что приводит к разрыву водородных связей между комплементарными нуклеотидами и расхождение нитей ДНК.
2. Элонгация - это передвижение РНК-полимеразы вдоль нити ДНК, сопровождающееся образованием фосфодиэфирных связей между рибонуклеотидами. Присоединение рибонуклеотидов происходит в соответствии с принципом комплементарности.
Этот синтез идет от 5’ конца к 3’ концу со скоростью 40- 50 нуклеотидов в секунду. Данная фаза протекает до тех пор, пока ДНК-полимераза не достигнет стоп-сигнала, после чего происходит терминация. В результате процессов транскрипции образуется транскрипт (пре-иРНК). Он почти полностью соответствует транскриптону.
Начинается следующий этап - процессинг - посттранскриптационное созревание РНК. Заключается в:
- удаление излишков - вырезаются неинформативные участки;
- сплайсинг - сшивка, соединение информативных участков. При этом иРНК укорачивается. Из пре-иРНК образуется иРНК. Далее иРНК соединяется с белком-информером, в результате чего образуется комплекс иРНК+ белок =информосома, который может выходить из ядра и транспортироваться в цитоплазму к рибосоме, где начинается следующий этап передачи информации – трансляция.
Центральный постулат генетики (Уотсон и Крик): ДНК®РНК®белок. В 70-е годы был обнаружен фермент – ревертаза (обратная транскриптаза), который позволяет по иРНК синтезировать участок ДНК (РНК®ДНК). Этот процесс называется обратной транскрипцией.
Трансляция (биосинтез белка)
Трансляция - перевод генетического текста иРНК в последовательность АК в белке.
Необходимый комплекс факторов:
1. строительный материал (20 АК);
2. молекулы тРНК (61 вид тРНК). 61 кодону соответствует 20 АК. 1 тРНК распознает более 1 кодона;
3. ферменты: аминоацил-тРНК-синтетаза - обеспечивают узнавание и связывание АК со своей тРНК (20);
4. молекула мРНК. У эукариот иРНК имеет так называемый "кэп" (от англ.- кепка, шапка) в области 5"-конца. Кэп представлен метил-ГТФ, необходимой для узнавания иРНК рибосомой и участвует в трансляции;
5. рибосомы, как место синтеза белка. Каждая рибосома состоит из малой и большой субъединиц. У прокариот они более мелкие - 70S: малая субъединица - 30S, большая - 50S. (S- скорость осаждения). У эукариот- 80S: малая - 40S, большая- 60S. Рибосома содержит рРНК, а также около 80 различных белков, в том числе и ферментов.
6. энергия АТФ и ГТФ;
7. ионы Мg2+, необходимые для активации ферментов;
8. белковые факторы трансляции – специфические белки различных этапов трансляции.
Этапы трансляции:
1. рекогниция – «распознавание»;
2. инициация – «начало»;
3. эллонгация – «продолжение, удлинение»;
4. терминация - «прекращение»;
5. процессинг – «созревание».
1. Рекогниция. Происходит связывание АК со своей тРНК: АК+тРНК®(над стрелкой – аминоацил-тРНК-синтетаза, АТФ) аминоацил-тРНК +АМФ+2Фн Количество этих реакций зависит от количества АК.
2. Инициация. Состоит из 7 фаз:
а) подготовка рибосомы к трансляции. Рибосома (30S) взаимодействует с фактором инициации 3 (ФИ-3), в результате разделяется на малую и большую субъединицы. Малая субъединица находится в комплексе с ФИ-3;
б) подготовка РНК-матрицы к трансляции. мРНК эукариот в области 5’-конца имеет особую структуру – КЭП, представленную метил-ГТФ. В этой фазе происходит присоединение к КЭПу КЭП-связываемых белков. Они обозначают на мРНК место прикрепления рибосомы;
в) подготовка инициаторной аминоацил-тРНК. Кодон, с которого начинается считывание генетической информации, представлен триплетом АУГ – метионин. В качестве инициаторной будет выступать МЕТ-тРНК: МЕТ-тРНК+ГТФ+ФИ-2® инициаторный метионин-тРНК;
г) образование инициирующего комплекса. При этом инициаторная МЕТ-тРНК взаимодействует с малой субъединицей рибосомы с образованием инициирующего комплекса. Этому процессу способствует белковый ФИ-3, связанный с рибосомой;
д) связывание мРНК с инициирующим комплексом: инициирующий комплекс+ ФИ-1+ 5’-конец мРНК. При этом АТФ®АДФ+Фн;
е) поиск и комплементарное взаимодействие со стартовым кодоном. При этом происходит скольжение малой субъединицы рибосомы до момента обнаружения кодона АУГ и комплементарного взаимодействия с ним антикодона МЕТ-тРНК;
ж) формирование 80S-рибосомы. Большая субъединица присоединяется к комплексу, образующемуся на 6 стадии, что сопровождается затратой энергии ГТФ (ГТФ®ГДФ+Фн). Для этой реакции используется ГТФ, находящаяся в составе инициирующего комплекса. В образовавщейся рибосоме выделяют 2 участка:
- Р-участок, в котором находится МЕТ-тРНК. Здесь будет происходить образование пептидных связей – сайт Р;
- А-участок (аминоацильный участок) – служит для присоединения аминоацильной-тРНК.
В завершении этой фазы происходит высвобождение белковых факторов ФИ-1, ФИ-2, ФИ-3 и КЭП-связанных белков.
3. Элонгация. Состоит из 3-х фаз:
а. присоединение следующей аминоацил-тРНК в соответствии со смыслом следующего кодона матрицы. Процесс требует энергии ГТФ ®ГДФ + Фн+Q и белкового фактора - фактор элонгации-1 (ФЭ-1);
б. пептизация. Между АК-остатками образуются пептидные связи (между АК-1 и АК-2). Данную реакцию катализирует пептидил-трансфераза. Этот фермент разрушает связь между первой АК и ее тРНК. Первая тРНК покидает рибосому. В результате в амино-ацильном участке рибосомы находится растущий пептид, а Р-участок освобождается;
в. транслокация или перемещение. Рибосома перемещается на 1 кодон в направлении 3’-конца тРНК. При этом пептдтл-тРНК остается на месте и происходит внутририбосомный переход растущего пептида из А-участка рибосомы в Р-участок. Для данного процесса необходимы энергия ГТФ ®ГДФ + Фн+Q и белковый фактор - ФЭ-2. Основным результатом является освобождение А-участка рибосомы, в который может поступить следующая аминоацил-тРНК.
Процесс элонгации протекает циклически с 1 по 3 фазу до момента обнаружения стоп-кодона. При достижении рибосомой нонсенс-кодона элонгация прекращается и наступает терминация.
4. Терминация.
Нонсенс-кодон распознается белковыми R-факторами (факторы освобождения) в А-участке рибосом. В результате действия R-факторов обеспечивается диссоциация элементов трансляционного аппарата и в цитоплазму высвобождаются рибосома, иРНК, полипептид. СЭС между пептидом и тРНК гидролизуется с затратой энергии ГТФ ®ГДФ + Фн+Q. иРНК в дальнейшем может использоваться повторно для трансляции (до разрушения ее иРНКазами).
Собственно трансляция включает 3 этапа: инициация, элонгация, терминация.
5. Процессинг белка.
Это совокупность изменений в структуре белка (полипептида), которые заканчиваются формированием структурно и функционально зрелой белковой молекулы. Процессинг может быть 2-х видов:
- котрансляционный, протекающий во время трансляции;
- посттрансляционный – химическая можификация белка происходит после трансляции.
Включает несколько видов химической модификации:
- ограниченный протеолиз - это отщепление либо пептидного фрагмента, либо N-концевой АК (МЕТ);
- реакция ацетилирования - присоединение ацетильного остатка;
- фосфорилирование - присоединение остатка фосфорной кислоты с образованием сложных белков (фосфопротеины);
- гликозилирование АК с образование гликопротеинов и протеогликанов;
- гидроксилирование АК – присоединение –ОН группы. Наиболее часто данному процессу подвергаются ПРО и ЛИЗ с образованием гидроксипролина (ОПР) и гидроксилизина (ОЛИ). Этот процесс необходим при образовании коллагена;
- окисление АК;
- образование вторичной, третичной и четвертичной структур – характерно для олигомерных белков. При образовании данных структур белок сворачивается - процесс фолдинга. Для этого необходимы специализированные белки - шапероны. Они ускоряют сворачивание белков, исправляют некорректные формы вторичной, третичной структур. Может наблюдаться патология фолдинга: существуют т.н. отрицательные шапероны. Их присутствие в клетке приводит к неправильному фолдингу, что выражается в гибели клеток.
Адресование белковПосле синтеза и процессинга белки должны быть правильно размещены в клетке либо правильно выделены на экспорт. Этим управляют особые механизмы адресования белков.
В структуре белка имеется т.н. сигнальный участок, содержащий информацию о принадлежности данного белка к определенной органелле клетки или о выделение белка на экспорт. Функцию сигнального участка выполняет фрагмент аминокислотной последовательности либо углеводный компонент.
Принципы адресования заключаются в процессе распознавания белков определенными органеллами. При этом распознание сигнального участка происходит только в случае, если он «пришел» в правильное место.
Гидрофобные АК сигнального участка направляет белок в ЭПР (мембранная структура); гидрофильные АК - в жидкие среды клетки (цитоплазма) или во внутренне пространство органелл; углеводный компонент адресует белок в кровь или лимфу.
Регуляция биосинтеза белкаКлетки многоклеточного организма содержат одинаковый набор ДНК, но белки синтезируются разные. Например, соединительная ткань активно синтезирует коллаген, а в мышечных клетках такого белка нет. В эритроцитах содержится Нb, и информация о Нb содержится во всех клетках. С возрастом скорость синтеза изменяется.
Прокариоты.
Теория регуляции биосинтеза у прокариот была разработана в 1961 г. Ф. Жакобом и Ж. Моно. Основные положения теории:
1. неоднородность генетического материала. В геноме имеются:
- структурные гены, которые кодируют синтез структурных белков или ферментов;
- регуляторные гены. Обеспечивают регуляцию считывания информации со структурных генов.
2. регуляция биосинтеза происходит на этапе транскрипции;
3. регуляция осуществляется путем репрессии (подавление транскрипции) и индукции (разрешение транскрипции).
Пр.: работа лактозного оперона.
В клетках микробов имеется фермент лактаза, участвующий в расщеплении лактозы до галактозы и глюкозы. Этот фермент в их клетках вырабатывается только при наличии субстрата – лактозы. Регуляция осуществляется с помощью лактозного оперона. Так, если в среде нет лактозы, то ген-I кодирует синтез белка-репрессора. Белок-репрессор взаимодействует с оператором и в результате РНК-полимераза не может осуществлять транскрипцию. Этот тип регуляции называется репрессией.
Если в среде присутствует лактоза, то она препятствует взаимодействию белка-репрессора с операторм, РНК-полимераза считывает информацию и образуется мРНК лактазы. Т.о. лактоза является индуктором, т.е. веществом, препятствующим взаимодействию белка-репрессора с оператором, в результате чего усиливается и облегчается транскрипция гена лактазы.
У прокариот первичные транскрипты генов (мРНК) используются в синтезе белка еще до завершения процесса транскрипции. У них нет ядерной мембраны.
Эукариоты.
Основные уровни регуляции биосинтеза:
1. на уровне транскрипции. Варианты:
- групповая репрессия генов белками – гистонами;
- амплификация генов - увеличение числа копий заданного участка ДНК или гена. Достигается в результате многократного синтеза ДНК в одном и том же репликативном пузыре. В этом случае транскрипция будет возможна сразу с нескольких копий гена, что увеличивается скорость транскрипции. Эта регуляция изучается у опухолевых клеток, которые способны к амлификации;
- регуляция сигналами-усилителями. Сигналы-усилители - энхансеры - выступающий участок ДНК, который может быть значительно удален от промотора. Под действием энхансера наблюдается более чем 200-кратное увеличение скорости транскрипции. Действует неспецифично, усиливая транскрипцию многих генов.
Пр.: действие гормонов коры надпочечников: глюкокортикоиды проникают внутрь клетки, где взаимодействуют с рецептором, посредством чего проникают в ядро, где присоединяются к ДНК и превращают участок ДНК в энхансер. При этом запускается синтез ферментов, характерных для действия глюкокортикоидов. Данный механизм работает только у эукариот.
2. регуляция на уровне процессинга иРНК:
- разрешение или запрещение процессинга. Так, не все пре-иРНК превращаются в зрелые иРНК: пре-иРНК ®иРНК;
- дифференциальный (альтернативный) процессинг. В клетках эукариот возможен многовариантный процессинг, поэтому утверждение 1 ген®1 белок для них не всегда справедливо. Это происходит в результате потери некоторых экзонов.
Пр.: С-клетки щитовидной железы и нейроны имеют одинаковый ген, который в С-клетках кодирует выработку кальцитонина (регулирует уровень Са2+), а в нейронах дифференцирует процессинг белка CGRP-пептид (регулирует АД).
3. на уровне стабильности и активности иРНК. Чем больше иРНК находится в стабильном состоянии в цитоплазме, тем большее количество молекул белка на ней может быть синтезировано. Поэтому в цитоплазме иРНК консервируются путем взаимодействия с белками-информатионами, образуя комплексы - информосомы.
4. регуляция на уровне трансляции:
- тотальная репрессия или активация трансляции при изменении активности и количества белковых факторов (ФИ и ФЭ);
- избирательная дискриминация иРНК, например, при инфицировании клетки вирусом транслируется вирусная РНК, а РНК хозяина дискриминируется.
- механизм повышения эффективности трансляции включает образование полисом - это комплекс нескольких рибосом с одной иРНК. Расстояние между соседними рибосомами составляет 80 нуклеотидов.
Нарушения матричных биосинтезов
ДНК человека содержит около 3 млрд. нуклеотидов. Точность их копирования зависит от точности: а) репликации, б) транскрипции, в) трансляции.
Факторы нарушения структуры ДНК: УФО, ионизация, химические агенты, спонтанные изменения (при фоновых излучениях).
Чаще всего происходит депуринизация (50"000 нуклеотидов за 70 лет жизни) – 40% всех пуриновых нуклеотидов (за 70 лет). Реже – дезаминирование и депиримидирование.
Этим процессам препятствуют процессы репарации ДНК. Т.о., постоянство информации поддерживается с помощью репликации и репарации.
Изменчивость генотипа наблюдается в результате мутаций. Их молекулярной основой является нерепаративное изменение первичной структуры ДНК. Напр., при действии азотной к-ты изменяются Ц→У, Ц→Г, У→А – эти изменения репаративная система "не замечает".
Система репарации ДНК
Репаративная система состоит из трех ферментов: (1) эндонуклеаза, (2) экзонуклеаза, (3) ДНК-полимераза (репарирующая).
Механизм репарации ДНК:
1. ДНК-эндонуклеаза обнаруживает участок повреждения и вызывает в этом месте разрыв фосфодиэфирных связей.
2. ДНК-экзонуклеаза отщепляет с образовавшихся концов нуклеотиды, в том числе и поврежденные.
3. ДНК-полимераза репарирующая восстанавливает нуклеотиды по принципу комплементарности.
Генные мутации
Они затрагивают небольшие участки ДНК (на уровне генов). Виды генных мутаций:
1. Замена нуклеотидов или кодонов. Замена одного нуклеотида – точечная мутация.
1.1. Замена нуклеотида без изменения смысла кодона. Напр., ААА (лиз)→ ААГ (лиз)
1.2. Замена нуклеотида с изменением смысла кодона. Напр., ААГ (лиз)→ ГАГ (глу). Это миссенс-мутация.
1.3. Замена с образованием терминирующего кодона. Напр., ААА (лиз)→ УАА (обрыв транскрипции). Это нонсенс-мутация.
2. Вставка.
2.1. Вставка одного или нескольких кодонов (т.е. триплетов), при этом не происходит сдвига рамки считывания. Напр., ААА ГЦА ГГА ЦЦА → ААА ГАГ ГЦА ГГА ЦЦА. В белке появляется одна или несколько "лишних" АК.
2.2. Вставка 1, 2 и др., но не кратного трем кол-ва нуклеотидов. Происходит сдвиг рамки считывания. Напр., ААА ГЦА ГГА ЦЦА → ААА ГАГ ЦАГ ГАЦ ЦА… Получается пептид со случайной аминокислотной последовательностью.
3.Делеция.
3.1. Выпадение одного или нескольких кодонов. Нет сдвига рамки считывания. Образуется белок, укороченный на одну или несколько АК.
3.2. Выпадение нуклеотидов 1, 2, но не кратного трем кол-ва. Сдвиг рамки считывания. Синтезируется пептид со случайной последовательностью АК.
Мутации, возникающие в половых клетках, передаются по наследству. Мутации соматических клеток могут приводить к раковому преобразованию клетки.
По биологическим последствиям мутации:
ü молчающие – не проявляются в обычных физиологических условиях, а проявляются в экстремальных условиях.
ü нейтральные – формируется нормальный белок (напр., при замене ГЛУ на АСП).
ü вредные (патогенные) – приводят к заболеванию (напр., при замене в гемоглобине в 6-м положении АК ГЛУ на ВАЛ развивается серповидно-клеточная анемия.
ü полезные – способствуют лучшей адаптации.
Ингибиторы матричных биосинтезов (Антибиотики)
а) ингибиторы, модифицирующие матрицы,
б) модифицирующие рибосомы,
в) инактивирующие ферменты.
Ингибиторы трансляции применяются против прокариот (это антибактериальные препараты). К ингибиторам транскрипции и репликации чувствительны и прокариоты, и эукариоты – они используются для подавления злокачественного роста. Напр., флеомицин, связываясь с ДНК, подавляет репликацию; актиномицин Д ингибирует транскрипцию; митомицин образует ковалентные связи с ДНК (нарушает и транскрипцию, и репликацию). Они подавляют биосинтез во всех клетках.
Антибактериальные
Стрептомицин нарушает структуру рибосом.
Тетрациклин нарушает связывание аминоацил-тРНК с рибосомами.
Эритромицин нарушает образование пептидных связей, нарушает конформацию 50S-субъединицы рибосом.
Биохимический полиморфизм
В результате мутаций возникают различные варианты генов. Если эти варианты не летальны, то они наследуются. Так формируется генотипическая неоднородность (гетерогенность). Это ведет к фенотипической неоднородности. Следствием генотипической неоднородности является полиморфизм белков – это существование разных форм белка, выполняющих одинаковые функции, но отличающихся по физико-химическим свойствам (и по первичной структуре).
Это изобелки, в т.ч. изоферменты.
Так, известно более 300 вариантов гемоглобина, 30 вариантов глюкозо-6-фосфат-ДГ, 14 вариантов альфа1-антитрипсина, 30 вариантов А,В-белков групп крови, 10 вариантов алкоголь-ДГ и ацетальдегид-ДГ.
Биохимический полиморфизм настолько велик, что в настоящее время говорят о биохимической индивидуальности.
Биохимический полиморфизм лежит в основе предрасположенности к заболеваниям (напр., атеросклерозу, алкоголизму, сахарному диабету), непереносимости некоторых пищевых компонентов (напр., лактозы – 15% европейцев, 80% азиатов), непереносимости лекарственных препаратов (напр., непереносимости препарата дитилина при сниженной активности холинэстеразы – дитилин используют при бронхиальной астме).
Биологические последствия обратной транскрипции
В молекулярной биологии принят основной постулат: ДНК → мРНК → белок. Однако, в 1970-е гг. Тёмин открыл фермент обратную транскриптазу (или ревертазу), катализирующую процесс обратной транскрипции (мРНК → кДНК - комплементарная ДНК). Этот процесс характерен для РНК-содержащих вирусов (ретровирусов), в которых имеется ревертаза, напр., для ВИЧ.
Обратная транскрипция заключается в синтезе короткого фрагмента ДНК, используя мРНК в качестве матрицы. Эта вирусная ДНК внедряется в ДНК хозяина (интеграция), что может приводить к образованию новых вирусных частиц. Интеграция беспорядочна и может привести (1) к мутациям, (2) к образованию онкогенов, (3) к усиленной работе эмбриональных генов (которые в норме во взрослом организме не функционируют).
Теломеры и теломеразы
Метод культуры клеток in vitro широко распространен. Если в эти условия поставить опухолевые клетки, то они делятся бесконечно долго – иммортализованные клетки. Нормальные клетки делятся ограниченное количество раз (это установил Л.Хэйфлик), есть предел деления – лимит Хэйфлика.
Напр., клетка новорожденного может делиться митозом 80-90 раз, взрослого – 60-70 раз, стариков – 20-30 раз, затем репликация нарушается и клетка гибнет.
В 1998 г. американские ученые смогли заставить нормальные клетки делить в 2 раза больше лимита Хэйфлика. В этом участвовали вещества теломеры и ферменты теломеразы.
Теломеры – это специализированные концевые районы хромосомной ДНК, состоящие из многократно повторяющихся коротких нуклеотидных последовательностей. Предполагалось, что эти вещества состоят из 6-8 нуклеотидов: TTAGGG – этот блок повторяется многократно, в результате длина их составляет 2-20 тысяч пар оснований.
Во время репликации в структуре ДНК имеется праймер – короткий РНК-олигонуклеотид, к которому присоединяется ДНК-полимераза. С этого места начинается репликация. Затем происходит удаление праймера и 5"-конец оказывается короче 3"-конца. Возникает эффект недорепликации (Оловников). Образовавшиеся нити ДНК наращиваются с помощью теломеразы. Теломераза синтезирует теломеры. Теломераза – РНК-содержащий фермент, она синтезирует по принципу обратной транскрипции концевой участок, а ДНК-полимераза наращивает другую цепь.
Т.к. теломераза синтезирует теломеры, то она повышает длительность жизни клетки. Повышенная активность теломераз делает клетку бессмертной. Активность теломеразы в различных клетках различна. По активности теломераз выделяют:
1. Соматические клетки. Теломеры их содержат 10-12 тысяч пар нуклеотидов и отсутствует теломеразная активность. Но в макрофагах и лейкоцитах теломераза активна.
2. Половые клетки. Теломеразная активность высокая, теломеры содержат больше – 15-20 тысяч –пар нуклеотидов. То же и в стволовых клетках.
3. Раковые клетки. Длина теломер не очень большая (8-10 тысяч пар нуклеотидов), но высокая теломеразная активность (у 80% раковых клеток).
Т.о., современная биохимия имеет задачей:
а) увеличение теломеразной активности в отдельных клетках, напр., в клетках кожи для восстановления ее после ожогов;
б) снижение теломеразной активности в раковых клетках.
Патология белкового обмена
Нарушения обмена белков могут наблюдаться на различных этапах:
1. На этапе поступления питательных белков в организм. В сутки человеку требуется около 100 г белка. Т.к. белки содержат незаменимые АК, то недостаточное поступление их ведет к снижению или даже к отсутствию синтеза белков в организме.
Напр., недостаток:
ЛИЗ – тошнота, головокружение, повышенная чувствительность к шуму; недостаток
ТРИ – снижение массы тела, гипопротеинемия;
ГИС – снижение гемоглобина в крови;
МЕТ – развитие жирового перерождения печени и почек.
Общее снижение кол-ва белка → белковая недостаточность → отрицательный азотистый баланс, гипопротеинемия.
Тяжелая форма белковой недостаточности – квашиоркор. Сначала снижается общий белок, снижение альбумина ведет к отекам (из-за изменения онкотического давления), снижение гемоглобина ведет к анемии, а само снижение синтеза белка приводит к гипераминоацидемии (повышение АК в крови) и к аминоацидурии. Также снижается синтез ферментов поджелудочной железой (трипсин, химотрипсин, полипептидазы – тоже белки), что ведет к снижению усвоения белка в кишечнике.
2. Нарушение на этапе пищеварения.
2.1. В желудке. Возможна гипоацидитас и анацидитас – ахлоргидрия (снижение и отсутствие кислотности в желудке соответственно). При ахлоргидрии начинается гниение белков.
Гиперацидитас – усвоение белка не нарушается, но может быть поражение слизистой желудка, переходящее в язву.
2.2. В тонкой кишке. Нарушение усвоения белка при панкреатитах, снижении секреции трипсина, химотрипсина.
2.3. В толстой кишке. Повышение процессов гниения белка, напр., при запорах, непроходимости кишечника.
3. Нарушение обмена белков в тканях, т.е. на уровне межуточного обмена. Может быть связано нарушением обмена АК.
3.1. Приобретенные нарушения обмена связаны с дефицитом витаминов; особенно В6 – нарушение процессов переаминирования и дезаминирования АК, развивается аминоацидурия. Либо связаны с гормональными нарушениями обмена АК.
3.2. Наследственные нарушения обмена АК.
Пример 1: в норме фенилаланин (ФЕН) под действием фенилаланингидроксилазы (ФАГ) окисляется кислородом до тирозина (ТИР).

При наследственной патологии (нарушении образования ФАГ) ФЕН накапливается в тканях и затем превращается в фенилпируват, который может превращаться в фениллактат или в фенилацетат. Они накапливаются в тканях и выделяются с мочой (фенилкетонурия). Эти соединения токсичны для ткани мозга, их накопление вызывает нарушения физического и умственного развития. При дефиците ФАГ развивается фенилпировиноградная олигофрения. Если она развилась по гомозиготному типу, то ребенок умственно отсталый и нуждается в помещение в специализированное учреждение. Необходима ранняя диагностика фенилкетонурии (первые 7-10 дней после рождения). Если ребенку ставится этот диагноз, то ему назначается диета, обедненная фенилаланином. Диета сохраняется до 16-18 лет (средний уровень развития).
Пример 2: Генетические нарушения обмена тирозина (ТИР). В организме ТИР образуется из ФЕН (катализируется ФАГ). Затем ТИР может превращаться: (1) в меланин, (2) в тиреоидные гормоны, (3) в ДОФА, а затем в адреналин, (4) в гомогентизиновую к-ту, а затем в конечные продукты (в мочу). Если нарушается блок "ТИР → меланин" (катализируется тирозиназой), то наблюдается альбинизм (отсутствие пигмента кожи меланина). Если блок "гомогентизиновая к-та → конечные продукты" (катализируется оксидазой в присутствии аскорбиновой к-ты), то наблюдается алкаптонурия: моча приобретает темно-бурую окраску, вплоть до черной). Также алкаптонурия может быть приобретенной – при авитаминозе С.



Пример 3: Гистидинемия – повышение ГИС в крови. В норме ГИС под действием гистидазы превращается в уроканиновую к-ту (5-формиминотетрагидрофолиевую к-ту). Накапливающийся ГИС приводит к нарушению умственного и физического развития.
4. Нарушения на стадии биосинтеза белка. Чаще всего наблюдается усиленный синтез белка к.-л. клеткой (злокачественное новообразование).
Механизмы развития раковой опухоли
Рак – генетическое заболевание, т.е. повреждение генов.
Виды повреждений генов:
1) потеря гена,
2) собственно повреждение гена,
3) активация гена,
4) инактивация гена,
5) привнесение генов извне (внедрение добавочных генов).
В каждой клетке находится около 100"000 генов. В настоящее время признано, что ряд генов может превращаться в гены, несущие информацию о белках опухолевой клетки – онкогены. Часто это нормальные гены, но в норме функционирующие только в эмбриональном периоде или малоактивные, которые стали активными.
Механизмы активации генов представлены: а) мутацией генов и б) активацией генов.
В настоящее время выделено более 100 онкогенов. Помимо онкогенов есть еще антионкогены – гены-супрессоры опухолей (ГСО). Белковые продукты этих генов препятствуют превращению нормальной клетки в опухолевую. Наиболее часто онкогены и ПСО встречаются среди генов белков-передатчиков сигналов регуляции роста и размножения клеток.
Онкогены образуются:
1) среди генов факторов роста,
2) среди R факторов роста,
3) среди белков-передатчиков сигнала внутрь клетки,
4) среди факторов транскрипции,
5) среди генов белков, связанных с ростом и размножением клетки.
Активация одного онкогена или потеря функций одного гена-супрессора недостаточна для превращения нормальной клетки в опухолевую; подсчитано, что клетке человека необходимо для этого 10 мутаций. Но выявлены гены, появление которых способствует опухолевому росту, так перерождение гена Р53 – в 50%. Ген Р53 кодирует информацию о белке клеточного роста. Это ген апоптоза – ген, приводящий к гибели поврежденную клетку. Если этот ген поврежден, то сохраняются клетки с мутационными изменениями, что приводит к накоплению мутаций. Это объясняет возникновение опухолевого роста у пожилых людей.
Считается, что вирусы вносят в клетку новые онкогены, которые повышают вероятность развития опухоли. Также число мутаций повышают физические и химические факторы.
5. Нарушение обмена белка на стадии образования конечных продуктов.
5.1. Чаще всего нарушения в орнитиновом цикле мочевинообразования. При этом нарушения на разных уровнях приводят к накоплению разных продуктов (предшествующих нарушенному процессу). Напр., нарушение образования карбомаилфосфата ведет к гипераминоацидемии (повышению АК в крови), накоплению аммиака. Либо нарушение аргининосукцинатсинтазной реакции [рисунок: цитруллин + АСП → аргининосукцинат, над стрелочкой "аргининосукцинатсинтаза" ] ведет к накоплению цитруллина.
Все нарушения орнитинового цикла приводят к слабоумию (напр., аргининосукцинатное слабоумие).
5.2. Нарушение выделения продуктов белкового обмена. Напр., цистиноз – нарушение реабсорбции АК (особенно цистина и цистеина) – цистинурия (в 20-30 раз ! ).
Генная инженерия
1970 г. – американский ученый Тёмин открыл фермент ревертазу (обратную транскриптазу), которая катализирует синтез РНК → кДНК (комплементарная ДНК, не содержащая интронов).
Генная инженерия – это искусственное создание активных генетических структур (рекомбинантных ДНК). Первая работа – 1972 г. – амер. Пол Берт.
Предпосылки генной инженерии
1) изучение (в деталях) матричных биосинтезов;
2) установление возможности обмена между двумя молекулами ДНК (т.е. рекомбинанции);
3) открытие ферментов рестриктаз (1970), разрезающих ДНК в определенных участках с образованием фрагментов с "липкими концами";
4) открытие ферментов ревертаз (1970);
5) обнаружение маленьких кольцевых ДНК в плазмидах.
Основные этапы генной инженерии:
1. получение необходимого гена:
а) с помощью рестриктаз "нарезается" ДНК, затем выделяется нужный фрагмент;
б) с помощью ревертаз по имеющейся мРНК синтезируется комплементарная ДНК (кДНК).
2. подготовка вектора – плазмиды или фага. [рисунок: кольцевая ДНК (плазмида) → (над стрелкой "рестриктаза") плазмида с вырезанным кусочком]
3. соединение к вектору необходимого гена [рисунок: большой отрезок плазмиды из предыдущего рисунка плюс кусок ДНК из первого рисунка → рекомбинантная ДНК кольцевого строения, над стрелкой "лигаза" ]
4. вектор внедряется в кишечную палочку (E.coli), клонирование гена (т.е. получение его копий).
Обмен углеводовУглеводами называются альдегиды или кетоны многоатомных спиртов или их производных.
Углеводы классифицируются на:
1. моносахариды – не подвергаются гидролизу:
- триозы (глицеральдегид, диоксиацетон);
- тетрозы (эритроза);
- пентозы (рибоза, дезоксирибоза, рибулоза, ксилуоза);
- гексозы (глюкоза, фруктоза, галактоза).
2. олигосахариды – состоят из 2–12 моносахаридов, соединенных между собой гликозидными связями (мальтоза – 2 глюкозы, лактоза – галактоза и глюкоза, сахароза – глюкоза и фруктоза);
3. полисахариды:
- гомополисахариды (крахмал, гликоген, клетчатка);
- гетерополисахариды (сиаловая кислота, нейраминовая кислота, гиалуроновая кислота, хондроитинсерная кислота, гепарин).
Углеводы входят в состав клеток животных (до 2%) и растений (до 80%).
Биологическая роль:
1. энергитическая. На долю углеводов приходится около 70% всей калорийности. Суточная потребность для взрослого человека – 400-500 г. При окислении 1 г углеводов до воды и углекислого газа выделяется 4,1 ккал энергии;
2. структурная. Углеводы используются как пластический материал для образования структурно-функциональных компонентов клеток. К ним относятся пентозы нуклеиновых кислот, углеводы гликопротеинов, гетерополисахариды межклеточного вещества;
3. резервная. Могут откладываться про запас в печени, мышцах в виде гликогена;
4. защитная. Гликопротеины принимают участие в образовании антител. Гетерополисахариды участвуют в образовании вязких секретов (слизи), покрывающих слизистые оболочки ЖКТ, дыхательных и мочеполовых путей. Гиалуроновая кислота играет роль цементирующего вещества соединительной ткани, препятствующего проникновению чужеродных тел;
5. регуляторная. Некоторые гормоны – гликопротеины (гипофиза, щитовидной железы);
6. участвуют в процессах узнавания клеток (сиаловая и нейраминовая кислоты);
7. определяют группу крови, входя в состав оболочек эритроцитов;
8. участвуют в процессах свертываемости крови, входя в состав гликопротеинов крови, фибриногена и протромбина. Так же предупреждает свёртываемость крови, входя в состав гепарина.
Основным источником углеводов для организма служат углеводы пищи, главным образом крахмал, сахароза и лактоза.
Крахмал – это смесь двух гомополисахаридов: линейного (амилоза от 10% до 30%) и разветвленного (амилопектин от70% до 90%) строения. Крахмал содержится в основных продуктах питания: картофель до 10%, крупы – 70-80%.

Остатки глюкозы соединяются в амилозе и линейных цепях амилопектина с помощью a-1,4-гликозидных связей, а в точках ветвления амилопектина - с помощью a-1,6-гликозидных связей.
Крахмал, поступая с пищей в ротовую полость, после механической обработки будет подвергаться гидролизу с помощью a-амилазы слюны. Этот фермент является эндоамилазой, расщепляющей a-1,4-гликозидные связи. Оптимальный рН фермента находится в слабощелочной среде (рН=7-8). Поскольку пища в ротовой полости долго не находится, крахмал здесь подвергается лишь частичному гидролизу с образованием амилодекстринов.
Далее пища идёт в желудок. Слизистая оболочки желудка гликозидазы (ферменты, расщепляющие углеводы) не вырабатываются. В желудке среда резко кислая (рН=1,2-2,5) ,поэтому действие a-амилазы слюны прекращается, но в более глубоких слоях пищевого комка, куда не сразу проникает желудочный сок, действие фермента слюны продолжается и крахмал успевает пройти стадию гидролиза - эритродекстринов.
Основным местом переваривания крахмала служит тонкий кишечник. Здесь происходят наиболее важные стадии гидролиза крахмала. В двенадцатиперсной кишке, куда открывается проток поджелудочной железы, под действием ферментов ПЖЖ (a-амилазы, амило-1,6-гликозидазы и олиго-1,6-гликозидазы) будет идти гидролиз крахмала. Выделяющийся сок поджелудочной железы содержит бикарбонаты, которые участвуют в нейтрализации кислого желудочного содержимого. Образующийся при этом углекислый газ способствует перемешиванию пищевого комка, создаётся слабощелочная среда (рН=8-9). Образующиеся катионы натрия и калия способствуют активации панкреатических гидролаз (a-амилаза, амило-1,6-гликозидаза, олиго-1,6-гликозидаза). Эти ферменты завершают гидролитический разрыв внутри гликозидных связей, начатых a-амилазой слюны.
Эритродекстрины превращаются в ахродекстрины. Под влиянием a-амилазы панкреатического сока завершается разрыв внутренних a-1,4-гликозидных связей в крахмале с образованием мальтозы. a-1,6-гликозидные связи в точках ветвления гидролизуются под действием амило-1,6-гликозидазы и олиго-1,6-гликозидазы, которая является терминальной (последней) в этом процессе.
Т.о. три панкреатических фермента завершают гидролиз крахмала в кишечнике с образованием мальтоз. Из тех глюкозных остатков, которые в молекуле крахмала были соединены с помощью a-1,6-гликозидных связей, образовались дисахариды – изомальтозы.
Слизистая оболочка тонкой кишки (энтероциты) синтезирует мальтазы (изомальтазы), лактазы и сахаразы. Образующиеся в результате гидролиза мальтоза, изомальтоза являются временным продуктом гидролиза, и в клетках кишечника они быстро гидролизуются под влиянием кишечных мальтазы, изомальтазы на две молекулы глюкозы. Т.о. в результате гидролиза крахмала в органах пищеварения образуется конечный продукт – глюкоза.
В составе пищи кроме полисахаридов поступают и дисахариды (лактоза и сахароза), которые подвергаются гидролизу только в тонком кишечнике. В энтероцитах синтезируются специфические ферменты: лактаза и сахараза, которые осуществляют гидролиз этих дисахаридов с образованием глюкоз, галактоз и фруктоз. Продукты полностью перевариваются. Углеводы - моносахариды всасываются в кровь и на этом завершается начальный этап обмена углеводов в организме человека - пищеварение.
Было установлено, что для всасывания моносахаридов (глюкозы) в кровь необходимо наличие в энтероцитах:
- в цитоплазме - ионов калия, натрия, АТФ и воды.
- в биомембранах - специфических белков–переносчиков и фермента - АТФ-азы.
90% образовавшейся в результате гидролиза крахмала глюкозы всасывается в кровь и через систему воротной вены поступает в печень, где депонируется в виде резервного полисахарида - гликогена. Около 10% всасывающихся в кровь моносахаридов попадает в большой круг кровообращения, разносится к органам и тканям, которые используют их в метаболических реакциях.
С пищей в организм человека поступает клетчатка – полисахарид, состоящий из остатков b-D- глюкопиранозы. В ЖКТ человека она гидролизу не подвергается, поскольку не вырабатываются b-гликозидазы, которые расщепляют её до глюкозы.
Биологическая роль клетчатки:
1. формирует пищевой комок;
2. продвигаясь по ЖКТ она раздражает слизистую оболочку, усиливая секрецию пищеварительных желез;
3. усиливает перистальтику кишечника;
4. нормализует кишечную микрофлору.
Достигая толстого отдела кишки, она под действием ферментов микрофлоры подвергается частичному сбраживанию с образованием глюкозы, малата, газообразных веществ. Глюкозы образуется мало, но она всасывается в кровь.
Биологический синтез гликогена
Установлено, что гликоген образуется почти во всех клетках организма, однако наибольшее содержание гликогена обнаружено в печени (2-6%) и в мышцах (0,5-2%). Т.к. общая мышечная масса организма человека велика, то большая часть всего гликогена содержится в мышцах.
Глюкоза из крови легко поступает в клетки организма и в ткани, легко проникая через биологические мембраны. Инсулин обеспечивает проницаемость мембран, это единственный гормон, обеспечивающий транспорт глюкозы в клетки органов и тканей. Как только глюкоза поступает в клетку, она сразу же как бы запирается в ней. В результате первой метаболической реакции, катализируемой ферментом гексакиназой в присутствии АТФ, глюкоза превращается в фосфорный эфир – глюкозо-6-фосфат, для которого клеточная мембрана не проницаема. Глюкозо-6-фосфат теперь будет использоваться клеткой в метаболических реакциях (анаболизм, катаболизм). Из клетки глюкоза может обратно выйти в кровь только после гидролиза под действием фосфатазы (глюкозо-6-фосфатазы). Этот фермент есть в печени, почках, в эпителии кишечника, в других органах и тканях его нет, следовательно, проникновение глюкозы в клетки этих органов и тканей необратимо.
Процесс биосинтеза гликогена можно записать в виде 4-х стадий:
глюкоза® (гексакиназа, АТФ®АДФ) глюкозо-6-фосфат® (фосфоглюкомутаза) глюкозо-1-фосфат® (глюкозо-1-фосфат-уридин трансфераза) УДФ-глюкоза® (гликоген-синтетаза, +[C6Н10О5]n) [C6Н10О5]n+1 (это наращенный гликоген) +УДФ



Затем УДФ+АТФ®(нуклеозиддифосфаткиназа) УТФ+АДФ. Т.о. на присоединение 1 молекулы глюкозы тратмтся 2 молекулы АТФ.
Гликогенсинтаза является трансферазой, которая переносит остатки глюкозы, входящие в УДФ-глюкозу на гликозидную связь остаточного в клетке гликогена. При этом образуются a-1,4- гликозидные связи. Образование a-1,6-гликозидных связей в точках ветвления гликогена катализирует специальный фермент гликогенветвящий.
Гликоген в клетках печени накапливается во время пищеварения, и рассматривается как резервная форма глюкозы, которая используется в промежутках между приёмами пищи.
Распад гликогена
Он может идти двумя путями:
1. Основной – фосфоролитический - протекает в печени, почках, эпителии кишечника.
Схематически его можно записать в виде 3-х стадий:
а) [C6Н10О5]n (это гликоген)® (фосфорилаза А, +Н3РО4) глюкозо-1-фосфат +[C6Н10О5]n-1
б) глюкозо-1-фосфат® (фосфоглюкомутаза) глюкозо-6-фосфат
в) глюкозо-6-фосфат® (глюкозо-6-фосфатаза, +Н2О) глюкоза + Н3РО4
2. Не основной – амилолитический. его доля мала и незначительна. Протекает в клетках печени при участии:
- a-амилазы слюны, расщепляющей a-1,4-гликозидные связи;
- амило-1,6-гликозидазы, расщепляющей a-1,6-гликозидные связи в точках ветвления гликогена;
- g-амилазы, которая последовательно отрывает концевые остатки глюкозы от боковых цепей гликогена.
Гликогеновые болезни
Гликогеновые болезни - наследственные нарушения обмена гликогена, которые связаны с недостаточностью какого–либо фермента, участвующего в синтезе или распаде гликогена. Как правило, эта недостаточность выражена в снижении активности или полном отсутствии фермента.
Различают гликогенозы – болезни, связанные с нарушением процесса распада гликогена. При этом клетки печени, мышц, почек накапливают большое количество гликогена, что ведет к разрушению клеток. У больных наблюдается увеличение печени, гипоглюкоземия натощак, мышечная слабость. Обычно такие больные умирают в раннем возрасте. Наиболее часто встречаются следующие гликогенозы:
- болезнь Герса, связанная с недостаточной активностью или отсутствием фосфорилазы печени;
- болезнь Мак-Ардля, -//- фосфорилазы мышц;
- болезнь Помпе, -//- a-1,4-гликозидазы;
- болезнь Гори, -//- амило-1,6-гликозидазы;
- болезнь Гирке, -//- глюкозо-6-фосфатазы.
Агликогенозы – заболевания, которые характеризуются нарушением синтеза гликогена. У больных: гипогликемия натощак, судороги, рвота, потеря сознания, постоянное углеводное голодание мозга приводит к отставанию умственного развития. Больные погибают в раннем возрасте. Наиболее часто встречаются следующие агликогенозы:
- болезнь Льюиса, связанная с нарушением выработки или полным отсутствием гликогенсинтазы;
- болезнь Андерсена, -//- гликогенветвящего фермента.
Пути катаболизма глюкозы
В зависимости от функционального состояния клеток органов и тканей, они могут находиться в условиях достаточного снабжения кислородом или испытывать его недостаток, находиться в условиях гипоксии.
Следовательно, катаболизм глюкозы в организме можно рассматривать с двух позиций: в аэробных и анаэробных условиях.
Анаэробный путь распада глюкозы в тканях называется гликолизом, если в анаэробных условиях распадается глюкозный остаток гликогена, то этот процесс называется гликогенолизом. Оба эти процесса протекают в цитоплазме клеток. Конечным продуктом окисления будет являться молочная кислота. В процессе окисления будет выделяться энергия за счет реакций субстратного фосфорилирования. Основная биологическая роль – энергетическая. Окисление глюкозы и глюкозного остатка гликогена в тканях отличается только в начальных стадиях превращения, до образования глюкозо-6-фосфата. Схематически это можно представить как:
глюкоза® (гексакиназа, АТФ®АДФ) глюкозо-6-фосфат;
[C6Н10О5]n (это гликоген)® (фосфорилаза А, +Н3РО4) глюкозо-1-фосфат +[C6Н10О5]n-1
глюкозо-1-фосфат ® (фосфоглюкомутаза) глюкозо-6-фосфат
Основные стадии гликолиза и гликогенолиза:
Процесс гликолиза сложный и многоступенчатый. Условно его можно разделить на 2 стадии.
1 стадия – завершается образованием глицеральдегид-3-фосфата. 2 стадия – называется стадией гликолитической оксидоредукцией. Она сопряжена с образованием АТФ за счет реакций субстратного фосфорилирования, окислением глицеральдегид-3-фосфата и восстановлением пирувата в лактат.
1 стадия:
гексакиназа ↓ АТФ®АДФ
глюкозо-6-фосфатизомераза ↓

фосфофруктокиназа ↓ АТФ®АДФ

альдолаза ↓


Фосфодиоксиацетон под действием изомеразы может превращаться в глицеральдегид-3-фосфат.
2 стадия. На ней перед всеми формулами ставим 2, т.к. фосфодиоксиацетон изомеризовался и получилось 2 молекулы глицеральдегид-3-фосфата:
дегидрогеназа, +Н3РО4 ↓ НАД®НАДН2

дифосфоглицераткиназа ↓ АДФ®АТФ

фосфоглицеромутаза ↓

енолаза ↓

фосфоенолпируваткиназа ↓ АДФ®АТФ

ЛДГ ↓ НАДН2®НАД
Т.о. анаэробные превращения глюкозы в тканях завершается образование молочной кислоты. В процессе превращения глюкозы было израсходовано 2 молекулы АТФ для фосфорилирования глюкозы и фруктоза-6–фосфата (гексакиназная реакция и фосфофруктокиназная реакция).
С этапа образования триоз (альдолазная реакция) идет одновременная их окисление, в результате этих реакций образуется энергия в виде АТФ за счет реакций субстратного фосфорилирования (фосфоглицераткиназная и пируваткиназная реакции).
На этапе гликолитической оксидоредукции идет окисление гицеральдегид-3-фосфата в присутствии Н3РО4 и НАД-зависимой дегидрогеназы, которая при этом восстанавливается до НАДН2.
Митохондрии в анаэробных условиях блокированы, поэтому выделяемый в процессе окисления НАДН2 находиться в среде до тех пор, пока не образуется субстрат, способный принять его. ПВК принимает НАДН2 и восстанавливается с образованием лактата, завершая тем самым внутренний окислительно-восстановительный цикл гликолиза. НАД-окисленный выделяется и вновь может принимать участие в окислительном процессе, выполняя роль переносчика водорода.
Три реакции гликолиза являются необратимыми:
- гексакиназная реакция;
- фосфофруктокиназная реакция;
- пируваткиназная реакция.
Энергетический эффект гликолиза (гликогенолиза):
АТФ(глюкоза)=(2*2)–2=2
АТФ(гликоген)=(2*2)–1=3
Биологическая роль гликолиза – энергетическая. Гликолиз является единственным процессом в клетке, способным поставлять энергию в форме АТФ в бескислородных условиях. В кризисных ситуациях, когда клетки органов и тканей по каким то причинам находятся в анаэробных условиях, гликолиз является единственным источником скорой энергетической помощи для сохранения жизнедеятельности клеток, а в эритроцитах, где митохондрии отсутствуют, гликолиз вообще является единственным процессом, продуцирующим АТФ и поддерживающим их функции и целостность.
Гексозодифосфатный путь превращения углеводов в тканях
В аэробных условиях, когда в ткани в достаточном количестве поступает кислород, происходит подавление гликолиза. При этом уменьшается потребление глюкозы, блокируется образование лактата. Эффект подавления гликолиза дыханием получил название эффекта Пастера.
Глюкоза в аэробных условиях сгорает в клетках с образованием конечных продуктов - воды и углекислого газа. При окислении 1 моль глюкозы будет выделено 38 молекул АТФ, а при окислении 1 глюкозного остатка гликогена – 39 молекул.
Химизм реакций превращения глюкозы такой же, как и в аэробных условиях, но только до стадии образования пирувата.
Превращение глюкозы до пирувата протекает в цитоплазме, затем пируват поступает в митохондрии, где подвергается окислительному декарбоксилированию. Образовавшийся при этом АцКоА в дальнейшем окисляется в митохондриях с участием ферментов ЦТК и сопряженных с ними ферментов дыхательной цепи (ЦПЭ).
Реакция окислительного декарбоксилирования ПВК осуществляется при участии ряда ферментов и кофакторов:
1. дегидрогеназ (НАД, ФАД);
2. декарбоксилаз (ТПФ);
3. ацилтрансфераз (HS-KoA);
4. липоевой кислоты (ЛК), участвующей в переносе углекислого газа.
СН3-СО-СООН (это ПВК)® (пируватдегидрогеназа, НАД, ФАД, HS-KoA, ТПФ, ЛК) СО2 +НАДН2 +Н2О +3АТФ +СН3-С(О)-SKoA (это АцКоА, он поступает в ЦТК).
При окислении глюкозы в аэробных условиях энергия выделяется за счет реакций:
1. субстратного фосфорилирования на этапах превращения 1,3-дифосфоглицериновой кислоты, фосфоенол-ПВК, сукцинил-КоА;
2. за счет реакций окислительного фосфорилирования на этапах превращения глицеральдегид-3-фосфата, ПВК, изоцитрата, a-кетоглутаровой кислоты, сукцината, малата.
Энергетический эффект окисления:
АТФ (глюкозы)=2*(3+1+1+3+12)-2=38
АТФ (гликогена)=2*(3+1+1+3+12)-1=39
Конечные продукты образуются:
- углекислый газ на этапах превращения пирувата, оксалосукцината, a-кетоглутаровой;
- вода образуется на этапах превращения: глицеральдегид-3-фосфата, 2-фосфоглицериновой кислоты, пирувата, изоцитрата, a-кетоглутаровой кислоты, сукцината, малата.
Т.о. в отличие от анаэробного пути, аэробный путь окисления глюкозы является энергитически более эффективным и является основным путем обеспечения клеток энергией. При этом окисление идет с образованием конечных продуктов – углекислого газа и воды.
Гексозомонофосфатный путь превращения углеводов в тканях
Гексозомонофосфатный путь превращения углеводов в тканях (пентозофосфатный путь, апотолический путь) протекает в цитоплазме клеток органов и тканей и представлен двумя последовательными ветвями: окислительной и неокислительной.
Активность этого пути превращения глюкозы зависит от типа ткани и ее функционального состояния. Особенно активно глюкоза окисляется по этому пути в тканях и органах, где синтезируется много липидов: печень, кора надпочечников, жировая ткань, молочные железы. Биологическая роль этого пути связана, прежде всего, с производством 2-х веществ:
1. рибозо-5-фосфата и его производных, которые используется в клетках для биосинтеза важнейших биологических молекул: АТФ, ГТФ, HSKoA, НАД, ФАД и нуклеиновых кислот (ДНК, РНК);
2. НАДФ·Н2, которые в отличие от НАД·Н2 не окисляется в дыхательной цепи митохондрии, а используется как источник протонов и электронов для синтеза веществ, включающего реакции восстановления (ВЖК, холестерина, желчных кислот, стероидных гормонов, витамина D3). НАДФН2 используется для обезвреживания ядов и токсических веществ (в реакции связывания аммиака при восстановительном аминировании a-кетокислот).
Этот путь является единственным поставщиком пентоз для работающих клеток тканей и органов, и на 50% покрывает потребность в НАДФН2, следовательно основная биологическая роль этого пути – анаболическая.
Окислительная стадия пентозного пути превращения глюкоза отличается от классического гексозодифосфатного пути с этапа превращения глюкозы-6-фосфата и включает 5 реакций:
глюкозо-6-фосфат ®(глюкозо-6-фосфатдегидрогеназа, НАДФ®НАДФН2) 6-фосфоглюкозолактон ® (лактоназа, +Н2О) 6-фосфоглюконовая кислота® (дегидрогеназа 6-фосфоглюконовой кислоты, НАДФ®НАДФН2) 3-кето-6-фосфоглюконовая кислота ®(декарбоксилаза, -СО2) рибулозо-5-фосфат® (изомераза) рибозо-5-фосфат® (эпимераза) ксилуозо-5-фосфат
При определённых условиях на этом заканчивается окислительная стадия пентозного цикла. Между пентозами устанавливается подвижное равновесие: рибулозо-5-фосфат« (изомераза) рибозо-5-фосфат« (эпимераза) ксилуозо-5-фосфат
Однако в ряде случаев, когда в клетках отмечается дефицит кислорода, может протекать неокислительная стадия пентозного цикла. Основными реакциями этого этапа являются 2 транскетолазные реакции и одна трансальдолазная. Все они обратимы. В результате этих реакций образуются субстраты для гликолиза (фруктозо-6-фосфат и глицеральдегид-3-фосфат), а также вещества, характерные для пентозного пути превращения глюкозы. Схематически неокислительную стадию пентозного цикла можно записать так:
1. транскетолазные реакции:
а) ксилуозо-5-фосфат+рибозо-5-фосфа«(ТПФ) седогептулозо-7-фосфат+ глицеральдегид-3-фосфат;
б) ксилуозо-5-фосфат+эритрозо-4-фосфат«(ТПФ) фруктозо-6-фосфат+ глицеральдегид-3-фосфат;
2. трансальдолазная реакция:
судогептулозо-7-фосфат+ глицеральдегид-3-фосфат« фруктозо-6-фосфат+ эритрозо-4-фосфат
Баланс окислительной и неокислительной стадий гексозомонофосфатного пути превращения глюкозы в тканях можно записать в виде суммарного уравнения реакции:
6 глюкозо-6-фосфат+ 7Н2О+ 12НАДФ® 5 глюкозо-6-фосфат+ 6СО2 +12НАДФН2 +Фн
Глюконеогенез и другие источники глюкозы для организма человека
Глюкоза является основным углеводом крови. Её концентрация в течение суток колеблется в зависимости от энергозатрат и частоты приемов пищи, содержания углеводов в пище. Для взрослого человека содержание глюкозы в крови составляет от 3,3 до 5,5 ммоль/л. Поддерживается концентрация глюкозы в крови за счет процесса биосинтеза и распада гликогена, глюконеогенеза и за счет углеводов пищи.
Глюконеогенез - это процесс образования глюкозы из неуглеводных предшественников, которыми являются продукты распада белков, липидов и углеводов. Основными являются пируват, лактат. Промежуточными могут быть метаболиты ЦТК, а так же глицерин и АК. Ряд АК (АСП, ТИР, ФЕН, ТРЕ, ВАЛ, МЕТ, ИЛЕ, ГИС, ПРО, АРГ) тем или иным путем превращаются в метаболиты ЦТК – фумаровую кислоту, которая в дальнейшем превращается в ЩУК. Другие АК (ГЛИ, АЛА, ЦИС, СЕР) превращаются в пируват.
Глюконеогенез возможен не во всех тканях. Главным местом синтеза глюкозы является печень, в меньшей степени почки и слизистая оболочка кишечника.
Биологическая роль глюконеогенеза заключается не только в синтезе глюкозы, но и в возвращении лактата в клеточный фонд углеводов. За счет этого процесса поддерживается уровень глюкозы в крови при углеводном голодании и сахарном диабете. Этот путь является единственным, который поддерживает биоэнергетику жизненно важных тканей в кризисных ситуациях.
Большинство реакций глюконеогенеза представляют собой обратимые реакции гликолиза, за исключением 3-х, которые являются термодинамически необратимыми.:
1. гексакиназной;
2. фосфофруктокиназной;
3. пируваткиназной.
Эти реакции гликолиза имеют при глюконеогенезе обходные пути, которые связаны с образованием фосфоенолпирувата, фруктозо-6-фосфата и глюкозы.
Обходные реакции гликолиза:
Первая обходная реакция глюконеогенеза связана с образованием фосфоенолпирувата. Она протекает в 2 стадии. Сначала в результате реакции карбоксилирования пируват превращается в ЩУК. Эта реакция протекает в митохондриях, куда ПВК поступает из цитозоля. ЩУК в митохондриях восстанавливается в малат под действием МДГ (НАДН2). Мембраны митохондрий не проницаемы для ЩУК, малат же легко выходит в цитозоль, где окисляясь снова превращается в ЩУК. ЩУК в дальнейшем принимает участие в глюконеогенезе, вступая в реакции декарбоксилирования и фосфорилирования. Донором фосфатного остатка служит ГТФ, но может быть и АТФ.
а) CH3-CO-COOH (это ПВК)® (пируваткарбоксилаза (биотин), +СО2, +АТФ, +Н2О) СООН-СО-СН2-СООН (это ЩУК) +АДФ +Фн;
б) СООН-СО-СН2-СООН (это ЩУК)®(фосфоеноилпируваткарбоксикиназа, +ГТФ, +Н2О) СООН-С(О~РО3Н2)=СН2+ СО2 +ГДФ.
Вторая реакция связана с образованием фруктозо-6-фосфата:
фруктоза-1,6-дифосфат® (фосфатаза, +Н2О) фруктоза-6-фосфат+ Фн
Третья реакция связана с образованием глюкозы:
глюкозо-6-фосфат® (фосфатаза, +Н2О) глюкоза+ Фн
Образовавшаяся в процессе глюконеогенеза глюкоза может вновь поступать в клетки органов и тканей и принимать участие в метаболизме (использоваться в тканях как энергетический субстрат, откладываться про запас в виде гликогена, участвовать в анаболических реакциях).
В организме взрослого человека массой 70 кг, главным образом в печени, за сутки образуется около 80 гр. глюкозы.
Патология углеводного обмена
Нарушения углеводного обмена могут быть на различных этапах обмена веществ. Основными показателями нарушения является изменение концентрации глюкозы в крови (гипер-, гипоглюкоземия) и появление глюкозы в моче (глюкозурия). Концентрация глюкозы в крови взрослого здорового человека в норме составляет 3,3-5,5 ммоль/л. Появление глюкозы в моче возможно в случае превышения величины почечного порога, который для глюкозы составляет 10 ммоль/л.
Основными причинами развития нарушения углеводного обмена являются:
1. алиментарные. Употребление пищи, богатой углеводами, ведет к быстрому переполнению гликогенного резерва печени, мышц, развитию гиперглюкоземии, глюкозурии. При снижении двигательной активности происходит снижение окислительных процессов и усиление биосинтеза жиров в тканях, что ведет к развитию алиментарного ожирения;
2. при поражении слизистых оболочек ЖКТ. При этом в желудке нарушается образование HCl (гипохлоргидрия или ахлоргидрия), поступающие углеводы сбраживаются под влиянием ферментов микрофлоры с образованием лактата, а белки подвергаются гниению. Это создает благоприятные условия для развития микрофлоры и приводит к расстройству пищеварения в целом. При поражении слизистой тонкого кишечника нарушается гидролиз дисахаридов или всасывание продуктов гидролиза;
3. при поражении печени нарушается биосинтез и распад гликогена, глюконеогенез;
4. при поражении поджелудочной железы нарушается секреция ферментов (a-амилаз, олиго-1,6-гликозидаз), участвующих в гидролизе крахмала и гликогена.
Наиболее грозным заболеванием ПЖЖ является сахарный диабет. При этом поражаются В-клетки, они перестают вырабатывать гормон инсулин. Инсулин – единственный гормон, обеспечивающий транспорт глюкозы в клетки органов и тканей. В случае недостаточной его выработки или отсутствия вообще происходит нарушение биоэнергетики клеток, органов и тканей. В этом случае интенсивному окислению подвергаются белки и липиды, что сопровождается избыточной продукцией аммиака и Ац-КоА.
Для связывания токсичного аммиака отвлекаются кетокислоты (ЩУК и a-кетоглутаровая) из ЦТК, их концентрация резко падает, что приводит к снижению интенсивности окислительных процессов. ЦТК не в состоянии окислить все молекулы ацетил-КоА, образование которых увеличивается с усилением окисления белков и липидов. Создаются условия для их конденсации с образованием кетоновых тел. При сахарном диабете в крови наблюдается гиперкетонемия (норма - до 0,1 г/л) и кетонурия.
2СН3-СОSKoA (это ацетил-КоА) ®(Ац-КоА-трансфераза) ацетоацетил-КоА ® (деацилаза, +Н2О, -HS-KoA) ацетоуксусная кислота.
Ацетоуксусная кислота может превращаться в b-гидроксимасляную кислоту, при этом НАДН2®НАД. Также она может превращаться в ацетон с отщеплением СО2.




В норме содержание кетоновых тел в крови здорового человека до 0,1 г/л. При поражении печени нарушается процесс биосинтеза и распада гликогена, процессы глюконеогенеза.
Наследственные заболевания, как правило, связаны с нарушением синтеза ферментов, участвующих в метаболизме углеводов. Например, алактазия - неусвояемость углеводов молока (лактозы). Это связано с отсутствием фермента – лактазы, поэтому поступающие с молоком дисахариды не усваиваются. У детей проявляется в виде рвоты, тошноты, поноса, вздутия живота, происходит обезвоживание организма. Лечение: исключение лактозы из пищи и замещение на мальтозу, сахарозу, глюкозу.
Другая группа заболеваний может быть связана с наследственными нарушениями обмена гликогена:
1. гликогенозы, связанные с недостаточным количеством ферментов, участвующих в распаде гликогена (болезнь Гирке, Кори);
2. агликогенозы – заболевания, связанные с нарушением синтеза гликогена (болезнь Льюиса. Андерсона и т.д.).
Липиды
Липиды – это сложные органические вещества биологической природы, не растворимые в воде, но растворимые в органических растворителях.
Все липиды делятся на простые и сложные. Простые: триглицериды, стерины, стериды и воски. Сложные: фосфолипиды, гликолипиды. Фосфолипиды делятся на сфинголипиды и глицерофосфолипиды. К глицерофосфолипидам относятся: фосфатидилхолин, фосфатидилсерин, фосфатидилэтаноламин, фосфатидилинозит и плазмогены (ацетальфосфатиды). К гликолипидам: цереброзиды, ганглиозиды, сульфатиды.
Наряду с белками и углеводами, липиды являются основными продуктами питания. В организме человека они поступают с продуктами растительного и животного происхождения. Суточная потребность взрослого человека составляет 80-100 г. Липиды составляют 10-20% от массы тела. В среднем в теле взрослого человека содержится 10-12 кг. липидов. Из них 25% приходится на структурные липиды, остальные относятся к резервным. Установлено, что 98% резервных липидов находится в жировой ткани.
Резервные липиды (жиры) представлены триглицеридами (ТГ). Они используются для энергетических нужд организма. Важнейшими ТГ являются эфиры глицерина и ВЖК. ВЖК могут быть как предельными (пальмитиновая С15Н31СООН, стеариновая С17Н35СООН), так и непредельными (олеиновая С17Н33СООН, линолевая С17Н31СООН, линоленовая С17Н29СООН, арахидоновая С19Н31СООН).
Фосфолипиды (ФЛ), гликолипиды являются структурными компонентами биологических мембран клеток, они не имеют такой энергетической ценности, как ТГ. Они, как и стерины (холестерин - ХС) относятся к структурным липидам.
ХС является предшественником ряда биологически активных веществ (БАВ), например стероидных гормонов (эстрогенов, андрогенов, минерало- и глюкокортикоидов), витаминов группы D и желчных кислот. Арахидоновая кислота (С19Н31СООН), входя в состав ФЛ, может принимать участие в образовании гормоноподобных веществ (простагландинов, лейкотриенов).
Биологическая роль липидов:
1. структурная – входят в состав биомембран клеток (ФЛ, ГЛ, холестерин);
2. резервная – нейтральные жиры могут откладываться про запас в жировое депо;
3. энергетическая – при окислении 1 г липидов до воды и углекислого газа выделяется 9,3 ккал энергии. На долю липидов приходится примерно 50% всей калорийности;
4. механическая – входя в состав соединительной ткани, подкожной жировой клетчатки, липиды предохраняют внутренние органы от повреждения при механических травмах;
5. теплоизолирующая роль – входя в состав подкожной жировой клетчатки, липиды предохраняют органы от перегревания и переохлаждения;
6. транспортная – входя в состав биомембран клеток, липиды участвуют в транспорте веществ (катионов);
7. регуляторная – все стероидные гармоны являются липидами. Гармоноподобные вещества (простагландины и лейкотриены) образуются из липидов;
8. выполняет роль смазочного материала для кожи, предохраняют её от сухости и растрескивания;
9. участвуют в передаче нервных импульсов;
10. липиды являются основным источником эндогенной воды - при окислении 100 г липидов образуется 107 мл эндогенной воды (из 100 г углеводов 57 мл воды, а из 100 г белков – 41 мл воды);
11. растворяющая – желчные кислоты, являясь стеринами, участвуют в растворении жирорастворимых витаминов А, Д, Е и К;
12. питательная роль – с пищей в организм поступают незаменимые ВЖК, которые имеют 2 и более двойных связей.
Незаменимые ВЖК в организме не синтезируются, но их роль велика:
1. они являются обязательным структурным компонентом биомембран;
2. препятствуют всасыванию холестерина в кишечнике;
3. стимулируют синтез желчных кислот в печени;
4. тормозят образование ЛПОНП, предупреждая развитие атеросклероза;
5. понижают свёртывание крови и понижают возможность тромбообразования;
6. повышают защитные силы организма;
7. предупреждают развитие кожных заболеваний;
8. являются источником гормоноподобных веществ.
Богаты ненасыщенными ВЖК растительные масла, в которых их содержание составляет 50-55%. Для полного удовлетворения суточной потребности взрослому человеку достаточно получать 15-20 г этих масел.
Простагландины, простациклины, тромбоксаны и лейкотриеныЭто гормоноподобные вещества, которые в организме человека образуются из полиненасыщенных ВЖК. Главным предшественником этих соединений является арахидоновая кислота, которая в организме может образовываться из незаменимых ВЖК.
Установлено, что эти соединения могут образовываться во всех тканях и клетках организма за исключением эритроцитов. Они выполняют роль местных гормонов, вырабатываются в макроколичествах и регулируют функции клеток и тканей, находящихся по соседству или там же, где вырабатываются. Эти вещества имеют короткий период полураспада - живут от нескольких секунд до нескольких минут. После разрыва они подвергаются инактивации, накапливаются в печени и выводятся с мочой.
Биосинтез начинается с момента, когда под действием фосфолипазы А2 из ФЛ выделяется арахидоновая кислота, занимающая центральное место в метаболизме. В дальнейшем под влиянием фермента циклооксигеназы, а в лейкоцитах – липооксигеназы, в структуру арахидоновой кислоты включается молекула кислорода.
В случае образования простаноидов образуются циклические соединения, а лейкотриены являются ациклическими соединениями. простагландины и простациклины являются 5-членными циклическими соединениями, а тромбоксаны – 6-членными. [рис. схема образования, описанная выше]
Простациклины и простагландины обладают широким спектром действия. В эндокринных железах они стимулируют образование гормонов. В жировой ткани они тормозят липолиз. Действуя на одни и те же ткани, они порой оказывают противоположный эффект:
- регулируют сокращение гладких мышц бронхов, кишечника, матки;
- оказывают влияние на сокращение миокарда;
- регулируют кровоток в почках, контролируют выведение с мочой воды и электролитов;
- регулируют проницаемость капилляров, процессы тромбообразования.
В ЦНС простагландины раздражают центры терморегуляции, вызывая повышение температуры, лихорадку. Повышают чувствительность нервных окончаний к раздражающему действию гистамина.
Простациклин образуется главным образом в сердце и сосудах, препятствует образованию тромбов, способствует расширению сосудов, уменьшает артериальное давление.
Тромбоксаны образуются в тучных клетках и тромбоцитах. Они синтезируются в самом начале образования сосудистого тромба и запускают механизм, приводящий к образованию тромба, способствуя прилипанию тромбоцитов.
Лейкотриены образуются в лейкоцитах. Их биологический эффект связан с воспалительными процессами, иммунными и аллергическими реакциями. Они способствуют прилипанию лейкоцитов к стенке сосудов в местах воспаления. Также способствуют сокращению гладкой мускулатуры, регулируют тонус сосудов, особенно стимулируя сокращение коронарных артерий.
Переваривание липидовПоступая с пищей, липиды в ротовой полости подвергаются только механической обработке. Липолитические ферменты в ротовой полости не образуются. Переваривание липидов будет происходить в тех отделах ЖКТ, где будут создаваться условия для эмульгирования и гидролиза, где будет оптимальная реакция среды для ферментов.
Все эти условия у взрослого человека создаются в кишечнике. У детей первого года жизни слизистая оболочка желудка вырабатывает липазы. рН лежит в слабо кислой среде (рН=5,5). Под влиянием желудочной липазы расщепляются эмульгированные жиры молока. У взрослого человека хотя и вырабатывается желудочная липаза, но она не активна, поскольку рН желудочного сока в норме лежит в резко кислой среде (рН=1,5-2,5).
Переваривание жиров пищи начинается в тонком отделе кишечника, где создаются все условия для гидролиза. В переваривании участвуют:
- желчные кислоты, которые образуются в печени;
- бикарбонаты и ферменты поджелудочной железы;
- ферменты собственно слизистой оболочки желудка.
При поступлении пищи из желудка в двенадцатиперстную кишку, слизистой последней начинают выделятся регуляторы:
- химоденин;
- секретин;
- холецистокинин;
- энтерокинин.
Все они обеспечивают желчеобразование в печени, сокращение желчного пузыря, выделение панкреатического сока и стимуляцию секреции желез тонкого кишечника.
Также под влиянием бикарбонатов поджелудочной железы будет происходить нейтрализация соляной кислоты. При разложении образующейся угольной кислоты выделяется СО2, который способствует лучшему перемешиванию пищевого комка.
Основную роль в переваривании пищи играют желчные кислоты, которые образуются в печени из холестерина. В основе всех желчных кислот лежит структура циклопентанпергидрофенантрена:

Родоначальником желчных кислот является холевая кислота, которая содержит ОН-группу в 3, 7 и 12 положениях. Производными холевой кислоты являются:
1. хенодезоксихолевая кислота, которая имеет 2 ОН-группы в 3 и 7 положениях;
2. дезоксихолевая кислота, имеющая окси-группы в 3 и 12 положениях;
3. литохолевая кислота, имеющая одну окси-группу в 3 положении.
Большая часть желчных кислот в печени конъюгирована глицином и таурином. Их еще называют парные желчные кислоты, например гликохолиевые, таурохолиевые кислоты. Соотношение конъюгатов меняется в зависимости от характера принимаемой пищи. В случае преобладания углеводов увеличивается содержание глициновых конъюгатов. Если преобладают белки в диете, то увеличивается содержание тауриновых конъюгатов.
Выделение желчных кислот способствует:
1. эмульгированию жира;
2. активации панкреатических липаз, фосфолипаз;
3. способствуют всасыванию труднорастворимых в воде веществ: ВЖК, ХС, моношлицеридов, жирорастворимых витаминов.
При эмульгировании жир дробится на мелкие капельки, что значительно увеличивает поверхность контакта липида с ферментами. Желчные кислоты обволакивают эти капельки, препятствуя тем самым их слиянию. Таким образом стабилизируется эмульсия жира, который будет подвергаться гидролизу под влиянием панкреатических липаз. Помогают гидролизу жира ионы Са2+, которые образуют комплекс со свободными ВЖК. [рис. гидролиза трипальмитина: трипальмитин +3 воды® (липаза) глицерин+ 3 С15Н31СООН] [рис. гидролиза фосфатидилхолина: фосфатидилхолин +4 воды®(фосфолипазы А1, А2, С, D) глицерин + С15Н31СООН + С17Н31СООН +Н3РО4 +НО-СН2- СН2-Nº( СН3) 3] Фосфолипаза А1 действует на связь между глицерином и предельной ВЖК, А2 – между глицерином и непредельной ВЖК, С – между глицерином и фосфорной кислотой, D – между фосфорной кислотой и холином (HO-CH2-CH2-N+(CH3)3).
В результате гидролиза пищевого жира образуются спирты, фосфаты, ВЖК, азотистые основания, АК и другие соединения. Необходимо отметить, что в расщеплении жиров принимают участие и кишечные липазы, но их активность невысокая. К тому же они расщепляют только моноглицериды.
Установлено, что всасывание продуктов гидролиза жира имеет свои особенности. Легко всасываются простой диффузией в слизистую кишечника спирты, АК, фосфаты, короткоцепочечные ВЖК (10-14 атомов С), азотистые основания. Труднорастворимые в воде продукты гидролиза (ВЖК, моноглицериды, холестерин, жирорастворимые витамины) всасываются только в комплексе с желчными кислотами. Эти комплексы называются холеиновыми. В таком виде эти соединения проходят через биомембраны энтероцитов. В эпителии клеток ворсинок кишечника происходит распад холинового комплекса. При этом желчные кислоты сразу поступают в ток крови и через систему воротной вены попадают в печень, откуда они вновь поступают в составе желчи в желчный пузырь и могут вновь принимать участие в новом акте переваривания жира пищи.
Установлено, что общий фонд желчных кислот у взрослого человека составляет 2,8-3,5 г. При этом они совершают 5-6 оборотов в сутки за счёт печёночно-кишечной циркуляции. После того, как продукты гидролиза жира поступили в энтероциты, в стенке кишечника начинают синтезироваться жиры, специфичные для данного организма, которые по своему строению отличаются от пищевого жира.
Механизм ресинтеза жираРесинтез жира в стенке кишечника происходит следующим образом:
1. сначала продукты гидролиза (глицерин, ВЖК) активируются с использованием АТФ. Далее происходит последовательное ацилирование глицерина с образованием моно-, ди-, триацилглицеридов. Центральное место в синтезе жира в стенке кишечника занимает фосфатидная кислота. Из неё в сенке кишечника, как правило в первые часы после приема пищи, образуются простые (ТГ) и сложные (ФЛ) жиры. [рис. активации глицерина и ВЖК: глицерин® (глицеролкиназа, при этом АТФ в АДФ) a-глицерофосфат; R-CH2-CH2-COOH +HS-КоА® (ацил-S-КоА-синтетаза, при этом АТФ в АМФ)R-CH2-CH2-COSKoA]


2. [рис. присоединения к a-глицерофосфату активированного остатка ВЖК: a-глицерофосфат+ R1-COSKoA®(ацилтрансфераза) моноацилглицерин +HS-KoA®(ацилтрансфераза, +R2-CO-SKoA) диацилглицерин+ HS-KoA. Диацилглицерин может присоединять инозит с образованием фосфатидилинозита; серин – фосфатидилсерин (под действием ПФ отнимается СО2 и образуется фосфатидилэтаноламин, с прибавлением к которому 3 СН3 образуется фосфатидилхолин); также может присоединять воду с выделением фосфорной кислоты на место которой присоединяется 3 активированный остаток ВЖК]
Транспортные формы липидов в организмеЛипиды являются нерастворимыми в воде соединениями, поэтому для их переноса кровью необходимы специальные переносчики, растворимые в воде. Такими транспортными формами являются липопротеины плазмы крови, которые относятся к свободным липопротеинам (ЛП). Ресинтезированный жир в клетках кишечника, либо синтезированный жир в клетках других органов и тканей может быть транспортирован кровью только после включения в ЛП, где роль стабилизатора играют белки.
Мицеллы ЛП имеют наружный слой и ядро. Наружный слой состоит из белка, ФЛ и свободного ХС, которые имеют гидрофильные полярные группы и проявляют сродство к воде. Ядро формируется из ТГ и эфиров ХС. Все эти соединения, входящие в состав ядра, не обладают гидрофильными группами.
ЛП транспортируют: ФЛ, ТГ, холестерин. Могут транспортировать некоторые жирорастворимые витамины (А,D,Е,К). Выделяют 4 класса транспортных ЛП, которые отличаются друг от друга по химическому составу, размером мицелл и транспортируемым липидам. Поскольку они имеют разную плотность и скорость оседания в растворе NaCl, их разделяют на следующие группы:
ХМ – хиломикроны. Они образуются в стенке тонкого отдела кишечника;
ЛПОНП – липопротеины очень низкой плотности - образуются в стенке кишечника и печени;
ЛПНП – липопротеины низкой плотности - образуются в стенке кишечника, печени и эндотелии капилляров из ЛПОНП под действием липопротеидлипазы;
ЛПВП – липопротеины высокой плотности – образуются в стенке тонкой кишки и печени.
Таким образом, ЛП крови образуются и секретируются 2-мя видами клеток – энтероцитами и гепатоцитами. При электрофорезе белков сыворотки крови ЛП движутся в зоне a- и b-глобулинов, поэтому их по элекрофоретической подвижности могут обозначить как:
ЛПОНП – пре-b-ЛП
ЛПНП – b-ЛП
ЛПВП – a-ЛП
ХМ – как самые большие по размеру частиц и самые тяжёлые при электрофорезе не движутся и остаются на старте.
Принято считать, что ХМ отсутствуют в крови натощак, а синтезируются они в стенке тонкого кишечника особенно активно после приема жирной пищи. Они транспортируют в основном ТГ от клеток кишечника и жировых депо к клеткам органов и тканей. Имеют большие размеры мицелл и поэтому не проникают в стенки сосудов. Завершается распад ХМ через 10-12 часов после приема пищи под влиянием липопротеидлипазы печени, жировой ткани, эндотелия капилляров. Продукты гидролиза вовлекаются в клеточный метаболизм.
ЛПОНП и ЛПНП транспортируют преимущественно холестерин. Эти фракции приносят его в клетки органов и тканей, которые используют ХС для построения биомембран, для образования стероидных гормонов и витаминов группы D. Их ещё называют атерогенными фракциями (пре-b и b).
ЛПВП – осуществляют транспорт холестерина из клеток и тканей в печень, где он окисляется, превращаясь в желчные кислоты. Это антиатерогенная фракция.
ЛПОНП, ЛПНП и ЛПВП путём эндоцитоза поглощаются клетками печени, кишечника, почек, надпочечников, жировой ткани и разрушаются в лизосомах или микросомах.
Ресинтезированный жир в стенке кишечника соединяется с небольшим количеством белка и образует стабильные комплексные частицы, которые называются ХМ. Поскольку размеры частиц большие, то они не могут проникать из эндотелия клеток кишечника в кровяные капилляры. Они диффундируют в лимфотическую систему кишечника, а из неё в грудной проток и в кровеносное русло. Уже после приёма пищи через 1,5-2 часа начинают расти концевые ХМ, которые достигают максимума к 4-6 часу после приёма жирной пищи.
Активное поступление ХМ в печень, жировую ткань, где под влиянием ферментов липопртеидлипаз (регулируются гепарином) они распадаются с образованием глицерина и ВЖК. Часть ВЖК используется клетками, а часть с помощью транспортных белков крови. Завершается распад ХМ через 10-12 часов после приёма пищи.
Индекс атерогенности – соотношение ХС в ЛПОНП, ЛПНП и ЛПВП.
Катер = ( ХСлпнп + ХСлпонп ) / ХСлпвп. В норме индекс атерогенности 2-3, если же он выше 4, то очень велика вероятность развития атеросклероза.
Превращение липидов в тканяхВ тканях постоянно идут процессы распада и синтеза липидов. Основную массу липидов организма человека составляют ТГ, которые в клетке имеются в виде включений. Период обновления ТГ в разных тканях составляет в среднем от 2 до 18 суток. Распад липидов происходит в лизосомах, в меньшей степени в микросомах и цитоплазме клеток при участии тканевых липаз. В результате тканевого липолиза образуются общие метаболиты: глицерин и ВЖК, окисление которых до конечных продуктов (воды и углекислого газа) сопровождается выделением большого количества энергии в виде АТФ.
Окисление глицерина в тканях тесно связано с гликолизом и гексодифосфатным путём путем превращения глюкозы, в который вовлекаются метаболиты глицерина по следующей схеме:
киназа ↓АТФ ® АДФ
дегидрогеназа ↓ НАД ® НАДН2, Н2О, 3АТФ
дегидрогеназа, +Н3РО4 ↓ НАД ® НАДН2
дифосфоглицераткиназа ↓ АДФ®АТФ
фосфоглицеромутаза ↓
енолаза ↓
фосфоенолпируваткиназа ↓ АДФ®АТФ
+ HS-KoA, -СО2, |
ТПФ, липоевая кислота ↓ НАД®НАДН2

[каждая молекула НАД·Н2 идет в митохондрии, где дает 3 молекулы АТФ]
Энергетический эффект окисления глицерина: АТФ=(3+3+1+1+3+12)-1=22
Таким образом, энергетический эффект окисления 1 молекулы глицерина в кислородных условиях составляет 22 молекулы АТФ.
Окисление ВЖК в тканях изучал Кнопп (1904г), который показал, что процесс окисления ВЖК до конечных продуктов циклический. Он назвал окисление ВЖК b-окислением.
Все ВЖК, имеющие чётное количество углеродных звеньев в цепи, обязательно окисляясь, укорачиваются в результате 1 бета цикла на 2 углеродных звена и проходят последнюю стадию превращения масляной кислоты, которая имеет свои особенности.
Ленинджером в 1949 году было установлено, что окисление ВЖК идет в митохондриях. Линнен в 1954 году установил, что b-окисление в тканях сопровождается укорочением ВЖК на 2 углеродных звена. В дальнейшем Ац-КоА окисляется до конечных продуктов в ЦТК. Линнен описал все стадии окисления ВЖК. В наше время b-окисление ВЖК в тканях называется циклом Кноппа-Линнена.
Установлено, что процесс b-окисления начинается в цитоплазме клеток с активации ВЖК. Биомембраны митохондрий для ВЖК не проницаемы и их транспорт внутрь митохондрий возможен только при участии азотистого основания – карнитина. Ацил-КоА в цитоплазме соединяется с карнитином при участии трансферазы. Образовавшийся комплекс ацил-карнитин легко проникает в митохондрии. В межмембранном пространстве уже при участии митохондриальных трансфераз этот комплекс распадается. Карнитин возвращается в цитозоль, а ВЖК подвергается окислению в матрице. Т.о. карнитин выполняет роль челнока для ВЖК между цитоплазмой и митохондриями.
[рис. R-CH2-CH2-COOH (это ВЖК)® (Ацил-КоА-синтетаза, АТФ®АДФ+Фн, +HS-KoA) R-CH2-CH2-COSKoA (это ацил-КоА)®(дегидрогеназа, ФАД®ФАДН2, Н2О, 2АТФ) R-CH=CH-COSKoA (это еноилацил-КоА)®(еноилацилКоА-гидратаза, +Н2О) R-CH(ОН)-CH2-COSKoA (это b-гидроксиацил-КоА) ®(дегидрогеназа, НАД®НАДН2, Н2О, 3АТФ) R-C(O)-CH2-COSKoA (это b-кетоацил-КоА) ® (тиолаза, +HS-KoA) R-COSKoA (это ацил-КоА) + CH3-COSKoA (это ацетил-КоА, идет в ЦТК где выделяется 12 АТФ)]
Так идет окисление всех ВЖК. В результате одного b-цикла выделяется 5 молекул АТФ, АцКоА включается в ЦТК и дает еще 12 молекул АТФ. Т.о. в результате окисления 2-х углеродных атомов цепи ВЖК выделяется 17 молекул АТФ.
Окисление масляной кислоты.
Масляная кислота® бутирил-КоА® кротонил-КоА® b-гидроксибутирил-КоА® ацетоацетил-КоА. Особенность последней стадии окисления ВЖК в том, что в тиолазной реакции образуется 2 молекулы Ац-КоА, одна из которых не проходила начальный этап b-окисления с выделением 5 молекул АТФ.




АТФ(ВЖК)=n/2*17-6, где:
n – количество углеродных звеньев ВЖК;
17 - количество молекул АТФ, выделяющееся в результате 1 бета цикла окисления (12 – ЦТК, 5 – само b-окисление);
6 (1+5): 1 - та молекула АТФ, которая поглощается при активировании ВЖК;
5 – количество АТФ, которые не выделяются на последней стадиях окисления при превращении масляной кислоты.
Окисление ВЖК, имеющих нечётное количество углеродных звеньев.
Такие ВЖК поступают в организм человека с пищей морских организмов, растений, мяса жвачных животных и растений. Окисление происходит также, как и ВЖК, имеющих чётное число углеродных звеньев, но только до последней стадии, когда образуется пропионил-КоА.
CH3-CH2-COSKoA (это пропионил-КоА)® (карбоксилаза, биотин), +СО2, +Н2О, АТФ®АДФ+Фн) СООН-СН(СН3)-СOSKoA (это метитмалонил-КоА) ®(мутаза) СООН-СН2-СН2-СOSKoA (это сукцинил-КоА)
Образующийся сукцинил-КоА является одним из метаболитов ЦТК, куда он вовлекается для своего дальнейшего окисления.
ВЖК, окисляясь в клетке, дают большое количество энергии в виде макроэргических связей в молекулах АТФ, которая (энергия) используется мышцами (сердечной, скелетными), нервной тканью, при длительных физических нагрузках, в стрессовых ситуациях.
Биосинтез глицерина и ВЖК в тканяхБиосинтез глицерина в тканях тесно связан с метаболизмом глюкозы, которая в результате катаболизма проходит стадии образования триоз. Глицеральдегид–3–фосфат в цитоплазме под влиянием глицерол–дегидрогеназы (НАДН2) превращается в глицерол–3–фосфат, занимающий центральное место в биосинтезе простых и сложных жиров. Глицерол–3–фосфат, ацилируясь, превращается в фосфатидную кислоту, из которой образуются нейтральные и сложные жиры (ФЛ).
Биосинтез ВЖК в тканях происходит в эндоплазматической сети. Исходным материалом синтеза ВЖК в тканях является ацетил-КоА, который в цитозоль поступает из митохондрий, где образуется при окислении метаболитов белков, углеводного и липидного обменов. Также может образовываться из цитрата, который в силу ряда причин (при гиподинамии, избыточном питании), когда значительно снижается интенсивность окислительных процессов в ЦТК (в митохондриях) выходит в цитозоль, тем самым стимулируя синтез ВЖК. Цитрат в цитозоле под влиянием лиазы распадается на АцКоА и ЩУК. АцКоА участвует в биосинтезе ВЖК, а ЩУК при участии малатдегидрогеназы (НАДН2) восстанавливается до малата, который возвращается в митохондрии, где вновь окисляется до ЩУК. Тем самым завершается внутренний челночный цикл.
Условием для биосинтеза ВЖК в тканях:
- наличие ацетил-КоА;
- наличие НАДФН2, поступающего для реакции синтеза из пентозного цикла;
- наличие специальных белков-переносчиков (НS–АПБ-ацил переносящие белки);
- наличие энергии в форме АТФ; СО2; Н2О;
- наличие ферментов синтеза ВЖК.
Процесс биосинтеза ВЖК циклический. Каждый цикл включает 6 этапов. Ацетил-КоА используется только на 1 этапе, как “затравка синтеза” с образованием 3-х углеродного соединения малонил-КоА.
1 этап – образование малонил-КоА:
СН3-СОSKoA (это ацетил-КоА)® (АцКоА-карбоксилаза (биотин), +СО2, +Н2О, АТФ®АДФ+Фн) СOOН-СН2-СОSKoA (это малонил-КоА).
2 этап – перенос ацетила и малонила на специальные белки под действием соответствующих трансфераз:
СOOН-СН2-СОSKoA (это малонил-КоА) ® (малонилтрансфераза, +HS-АПБ, -HS-KoA) СOOН-СН2-СОSАПБ (это малонил-АПБ);
СН3-СОSKoA (это Ац-КоА)®(ацилтрансфераза, +HS-АПБ, -HS-KoA) СН3-СОSАПБ (это ацетил-АПБ).
3 этап – конденсация малонила и ацетила под влиянием синтетазы (конденсирующий фермент):
СOOН-СН2-СОSАПБ (это малонил-АПБ) + СН3-СОSАПБ (это ацетил-АПБ)®(конденсирующий фермент, -СО2, -HS-АПБ) СН3-С(О)-СН2-СОSАПБ (это ацетоацетил-АПБ).
4 этап – восстановление b-кетоацетила-АПБ при участии редуктазы (НАДФН2):
СН3-С(О)-СН2-СОSАПБ (это ацетоацетил-АПБ)® (редуктаза, НАДФН2®НАДФ) СН3-СН(ОН)-СН2-СОSАПБ (это b-гидроксибутирил-АПБ).
5 этап – дегидратация b-гидроксибутирила-АПБ:
СН3-СН(ОН)-СН2-СОSАПБ (это b-гидроксибутирил-АПБ)®(дегидратаза, -Н2О) СН3-СН=СН-СОSАПБ (это кротонил-АПБ или еноилацетил-АПБ).
6 этап – восстановление еноилацетила-АПБ при участии редуктазы (НАДФН2):
СН3-СН=СН-СОSАПБ (это кротонил-АПБ)®(редуктаза, НАДФН2®НАДФ) СН3-СН2-СН2-СОSАПБ (это бутирил-АПБ).
Т.о., в результате первого цикла синтеза ВЖК образуется 4-х углеродный фрагмент. Далее, в зависимости от того какую ВЖК в клетке нужно синтезировать, будут последовательно и циклично присоединяться молекулы малонил-КоА. Завершается синтез любой ВЖК реакцией гидролиза с участием фермента деацилазы: R-СН2-СН2-COSАПБ ®(деацилаза, +Н2О, -HS-АПБ) R-СН2-СН2-COОН (это ВЖК).
Для расчета количества молекул АТФ, необходимых для синтеза ВЖК, а также количества синтеза и количества молекул малонил-КоА можно воспользоваться формулой: n/2-1, где n – количество углеродных атомов ВЖК.
Таким образом, в организме можно синтезировать все предельные ВЖК и непредельные ВЖК, имеющие 1 двойную связь.
Биосинтез холестерина (ХС)

Основа - циклопентанпергидрофенантрен
Биосинтез ХС в клетках печени, тонкого кишечника и кожи осуществляется из АцКоА, который освобождается в результате распада белков, липидов и углеводов. Эндогенный ХС синтезируется преимущественно в печени (85%), в небольшой степени в тонком кишечнике (10%) и коже (5%).
Всего в организме взрослого здорового человека массой 70 кг за сутки образуется 2,8-3,5 г ХС. С пищей за сутки в организм поступает около 0,3 г ХС. Источником ХС для организма человека являются липиды пищи и эндогенный холестерин. Печень участвует в распределении ХС между клетками органов и тканей через транспортные ЛП крови. На экспорт ХС синтезируется только печенью и тонкой кишкой.
Холестерин является обязательным компонентом клеток. Наиболее богата ХС плазматическая мембрана гепатоцитов, где на его долю приходится 30% всех мембранных липидов. В миелине уровень ХС составляет 20%. В свободном виде и в виде эфиров он содержится в ЛП крови, входит в состав жировых включений цитоплазмы.
В организме человека ХС выполняет следующие функции:
1. структурная – ХС является структурным компонентом биомембран клеток. Принимает участие в регуляции их проницаемости, регулирует активность ферментов, работу рецепторов и переносчиков биомембран;
2. метаболическая – связана с тем, что ХС является предшественником стероидных гормонов (андрогены, эстрогены, минерало- и глюкокортикоиды), витамина D3 и желчных кислот.
Процесс биосинтеза ХС сложный и многоступенчатый и включает 35 реакций. Детали синтеза ХС практического врача не интересуют, поскольку многочисленные попытки управлять этим процессом в терапевтических целях оказались безуспешными, но знать схему биосинтеза надо. Источником БС ХС является ацетил-КоА, который образуется в результате катаболизма белков, углеводов и липидов.
2 ацетил-КоА (2 углеродных атома) ®(ацетил-КоА-тиолаза) ацетоацетил-КоА (4С) ®(гидроксиметилглутарил-КоА-синтетаза, +АцКоА) b-гидрокси-b-метилглутарил-КоА (6С) ®(редуктаза) 5 мевалоновая кислота (6С) ®(конденсация) сквален (30С) ®(циклизация) ланостерин (30С) ®(происходит деметилирование и перемещение двойной связи в кольце, -3СН3) холестерин (27С)
Освободившийся в результате распада клеток, ХС с помощью ЛПВП транспортируется в печень, где при участии цитохрома Р450 окисляется с образованием желчных кислот. примерно 80% окисленного ХС выводится с желчными кислотами через кишечник в виде капостеринов.
Патология липидного обменаНа этапе поступления с пищей. Обильная жирная пища на фоне гиподинамии ведёт к развитию алиментарного ожирения. Нарушение обмена может быть связано с недостаточным поступлением жира в организм с пищей, что приводит к развитию дерматитов, склероза сосудов из-за недостатка в незаменимых ВЖК. Полное исключение липидов из пищи ведет к авитаминозам (А, D, Е, К). Недостаточное поступление с пищей липотропных веществ ведет к нарушению синтеза ФЛ – структурных компонентов клеток – и в конечном итоге к развитию жировой инфильтрации печени, тканей (ФЛ®ТГ). Липотропными веществами являются: холин, инозит, серин – обязательные структурные компоненты ФЛ, а также витамин В6 (ПФ) - кофермент декарбоксилазы серина, метионин – донор метильных групп при биосинтезе фосфатидилхолина, фолиевая к-та и витамин В12 – коферменты метил-ТФ.
На этапе пищеварения. При поражении печени, желчных путей, желчного пузыря, когда в результате воспалительных процессов нарушается синтез или выведение в кишечник желчных кислот, когда нарушается биосинтез ЛП крови. Наиболее часто эти нарушения связаны с нарушением экскреция желчных кислот, что ведет к скоплению желчи в желчном пузыре, насыщении ее ХС. В этих условиях могут образовываться холестериновые камни. Застой желчи, воспалительные заболевания в печени и желчном пузыре способствуют осаждению ХС. Центрами образования холестериновых камней служат слущивающиеся клетки эпителия, на которые осаждается эпителий. Развитие желчекаменной болезни приводит к нарушению оттока желчи в кишечник и нарушению перевариванию липидов. В крови наблюдается гиперхолестеринэмия.
Поражение поджелудочной железы, слизистой тонкой кишки ведёт к нарушению переваривания жира. Развивается стеаторея, когда содержание жира в тканях увеличивается и составляет более 50%, кал становится ахоличный (преобретает серовато-белый цвет). Причины – нарушение образования или выведения желчных кислот. Нарушается функция поджелудочной железы (образование липаз, фосфолипаз, холестераз), воспаление слизистой в тонкой кишке, нарушается всасывание.
Липидный обмен нарушается при сахарном диабете. Накопление недоокисленных веществ ведёт к развитию кетоза. В случае недостаточной выработки инсулина интенсивному окислению подвергаются липиды клеток, что сопровождается избыточным Ацетил КоА, а это усиливает синтез ХС (гиперхолестеринэмия).
На этапе обмена ХС. Самым распространённым заболеванием является атеросклероз, которое характеризуется отложением ХС в стенке сосудов и тканей. Вокруг ХС формируются бляшки, развивается соединительная ткань (склероз), развивается кальцификация сосудов, тромбозы, нарушается кровоснабжение тканей. Болезнь развивается тогда, когда между клетками тканей и ЛП крови нарушается гомеостаз, развивается гиперлипопротеинэмия. Как правило, в крови растёт содержание фракций ЛПОНП и ЛПНП, которые снабжают клетку ХС. Снижение содержания ЛПВП – назначение которых удалять избыточный ХС. В клетки сосудов проникают все ЛП за исключением хиломикронов. Поскольку в их состав входят ФЛ, ТГ и белки, они быстро распадаются и удаляются из клетки. ХС выходит в межклеточное пространство. Он является чужеродным, поэтому его окружают марофаги. Все вещества, фагоцитирующие ЛП крови, разрушают ферменты лизосом за исключением ХС. Накапливаясь в большом количестве, откладывается в межклеточном пространстве, инкапсулируясь соединительной тканью, он ведёт к развитию склероза, к образованию атеросклеротических бляшек. Развитию его способствует курение, употребление алкоголя, сахарный диабет, обильная жирная пища и гипертоническая болезнь, т.е. все заболевания, которые ведут к повреждению стенок сосудов токсинами.
Гормоны
Гормоны - мобильные посредники в регуляторной системе. Термин «гормон» - от латинского «побуждать к действию».
Гормоны - это вещества, синтезирующиеся специальными железами, транспортируемые кровью и воздействующие на различные органы.
Гормоны характеризуются:
- синтезируются железами внутренней секреции;
- действуют дистантно;
- строгая специфичность действия;
- высокая биологическая активность (концентрация 10-12-10-15 моль/л).
В настоящее время установлено, что гормоны образуются во всех тканях.
Выделяют гормоны:
- с эндокринным эффектом - гормоны, синтезируемые ЖВС и поступающие в кровь;
- с паракринным эффектом - гормоны образуются в одном месте и действуют рядом;
- гормоны с аутокринным эффектом - действуют на ту ткань, где и образуются.
Для каждого гормона существует ткань-мишень. По первым представлениям, ткань-мишень - это та ткань, в которой гормоны вызывают физиологические или биохимические изменения. Например, тиреотропный гормон действует на щитовидную железу, следовательно, ткань-мишень - щитовидная железа; для инсулина - печень, мышечная ткань, жировая ткань.
По современным же представлениям, ткань-мишень - это ткань, в которой имеются рецепторы к данному гормону.
Концентрация гормонов значительно ниже, чем других БАВ, поэтому клетка-мишень должна отличить гормоны от других соединений, что осуществляется с помощью рецепторов - молекул узнавания. Связывание гормона с рецептором основано на комплиментарности какого-то участка мембраны. G+R®GR®G+R Комплекс «гормон-рецептор» может быть более активен, чем просто гормон.
Свойства рецепторов: связывают гормоны, генерируют сигналы (усиливают сигналы гормона).
Гормоны - это межклеточные регуляторы рецепторного действия (включая и гормоны и нейромедиаторы).
Классификация гормоновСуществует несколько видов классификации.
По месту образования гормонов:
1. гормоны гипоталамуса;
2. гормоны гипофиза;
3. гормоны щитовидной железы;
4. гормоны поджелудочной железы;
5. гормоны паращитовидных желез;
6. гормоны надпочечников;
7. гормоны половых желез;
8. гормоны местного действия.
По химическому строению:
1. белково-пептидные гормоны: гормоны гипоталамуса, гипофиза, поджелудочной железы, паращитовидных желез;
2. производные аминокислот: адреналин, норадреналин, тироксин, трийодтиронин;
3. стероиды: в их основе лежит структура циклопентанпергидрофенантрена, образуются из холестерина (половые гормоны, коры надпочечников).
По механизму действия (по расположению рецепторов):
1. гормоны, действующие через внутриклеточный рецептор - липофильные гормоны - стероиды и тиреоидные гормоны;
2. гормоны, действующие через рецепторы, находящиеся на поверхности клетки - гидрофильные гормоны. Они действуют через внутриклеточный посредник - мессенджер.
Гормон - первый посредник, а цАМФ, ионы Са2+, фосфатидилинозиды - вторые (чаще цАМФ, которая образуется из АДФ) посредники. [рис. цАМФ]
Механизм действия гормоновЛипофильные гормоны.
Гормон диффундирует через плазматическую мембрану и связывается внутренними рецепторами, образуется комплекс «гормон-рецептор», который активируется и действует на ДНК. G+R®GR®GR*®ДНК. В ДНК выделяют гормон-чувствительный элемент (ГЧЭ). Под его влиянием изменяется транскрипция, что влияет на деградацию мРНК. Гормоны влияют на процессинг белка. Гормоны действуют непосредственно на ДНК, активируют ферменты, стимулируя их синтез.
Гидрофильные гормоны.
Самый распространенный второй посредник - цАМФ-аденилатциклазная система. Она состоит из 2 частей: собственно аденилатциклазный компонент и протеинкиназный компонент.
В плазматической мембране находятся рецепторы 2 типов: Rs - стимулирующий и Ri - ингибирующий. Внутри мембран находится G-белок (Gs, Gi) (читается джи-белок). G-белок распадается на a и bg субъединицы, которые могут взаимодействовать между собой. a-субъединица и ГТФ действуют на аденилатциклазу, превращая ее в активную форму. Эта активная аденилатциклаза находится на внутренней поверхности ЦПМ. Под влиянием АЦ АТФ превращается в цАМФ+ФФн. Разрушается цАМФ с помощью фермента - фосфодиэстеразы.
После образования АЦ включается протеинциклазный компонент. цАМФ + протеинкиназа (фермент, вызывающий фосфорилирование других белков) ®активированая протеинкиназа. Протеинкиназа – тетрамер, содержащий 2 субъединицы R (регуляторная субъединица) и 2 субъединицы C (каталитическая субъединица). R2C2+4цАМФ«R2(4цАМФ) +2C (активная протеинкиназа). Активная протеинкиназа вызывает фосфорилирование белков.
Белок+ АТФ®(над стрелкой 2С) фосфопротеин (серин-фосфат, треонин-фосфат) +АДФ.
В результате образования фосфорилированных белков могут быть:
1. усиленный распад гликогена: фосфорилаза В®фосфорилаза А;
2. изменение транспорта ионов;
3. изменение метаболизма углеводов, липидов;
4. регулируется генная транскрипция.
Таким образом, фосфорилирование белков является важнейшим регуляторным механизмом.
Ионы Са2+.
Образуют соединение с белком - кальмодулин. Комплекс Са2+-кальмодулин активирует ферменты (аденилатциклазу, фосфодиэстеразу, Са2+-зависимую протеинкиназу). Есть группа гормонов, для которой второй посредник неизвестен - инсулин, гормон роста, пролактин.
Гормоны центральных желез - гипоталамуса и гипофизаРаздражитель действует на ЦНС, импульсы передаются в гипоталамус, который выделяет регулирующие факторы – либерины и статины. Эти факторы действуют на аденогипофиз, который выделяет тропные гормоны. Тропные гормоны действуют на периферические эндокринные железы, а те, в свою очередь, на ткани-мишени, в которых формируется физиологический или биохимический ответ. [можно оформить в виде вертикальной схемы со стрелочками] Также существует обратная связь между периферическими железами и тропными гормонами.
Гормоны гипоталамуса поступают в гипофиз через портальную систему сосудов.
Гормоны гипоталамуса:
1. кортикотропин - релизинг-гормон (кортиколиберин) – КРГ - влияет на секрецию АКТГ и выделение эндорфинов;
2. тиреотропин - релизинг-гормон (тиреолиберин) – ТРГ - влияет на секрецию ТТГ;
3. гонадотропин - релизинг-гормон - ГнРГ - влияет на секрецию лютеинизирующего (ЛГ) и фолликулстимулирующего (ФСГ) гормон;
4. гормон роста - релизинг-гормон - СТГ-РГ- влияет на секрецию гормона роста;
5. гормон, ингибирующий высвобождение гормона роста (соматостатин) – СС - тормозит секрецию гормона роста;
6. гормон, ингибирующий высвобождение пролактина (пролактинингибирующий гормон) – ПИГ - тормозит секрецию пролактина.
Многие гормоны гипоталамуса образуются и в других отделах НС и в периферических тканях, например, соматостатин – образуется в поджелудочной железе. Посредником в действии релизинг-факторов является цАМФ, но может использоваться и Са2+-фосфолипидный механизм. Гормоны гипоталамуса не только влияют на секрецию и выделение гормонов гипофиза, но и могут увеличивать и синтез гормонов.
Гормоны передней доли гипофиза (аденогипофиза):
1. простые белки;
2. гликопротеины;
3. пептиды семейства проопиомеланокортина (ПОМК).
Гормоны - простые белки:
1. гормон роста (ГР);
2. пролактин (ПРЛ);
3. плацентарный пролактин (лактоген) (ПЛ) - хорионический соматомаммотропин (ХС).
Эти гормоны стимулируют рост и лактацию. Содержат 190-195 аминокислот, они близки по строению и эффекту.
Гормон роста содержит 191 АК. Строение на 85% аналогично строению холестерина (ХС) и на 35%- с пролактином (ПРЛ). Концентрация гормона роста выше, чем других гормонов. Секретируется эпизодически. Влияют стрессовые состояния, белковая пища, физические упражнения – при этом повышается секреция.
Влияние на обмен веществ:
а) повышает синтез белка:
- повышает транспорт АК в клетки;
- повышает синтез РНК и ДНК;
- повышает интенсивность трансляции белков.
Гормон роста действует через инсулинподобный фактор роста (ИФР-1,2). ИФР-1- это главный посредник в действии гормона роста.
б) влияет на углеводный обмен - антагонист инсулина:
- повышает содержание глюкозы в крови - гипергликемия;
- снижает периферическую утилизацию;
- повышает продукцию глюкозы в процессе глюконеогенеза.
в) влияет на липидный обмен, стимулирует липолиз:
- увеличивает скорость окисления ВЖК (в печени).
г) влияет на минеральный обмен:
- задерживает в организме кальций, магний, фосфор, стимулирует рост костной ткани.
Гиперсекреция гормона роста у детей проявляется в виде гигантизма, у взрослых - акромегалия. Дефицит - гипосекреция - приводит к карликовости.
Пролактин - белок, состоящий из 198 АК. Его секреция увеличивается при беременности и лактации. Участвует в инициации лактации и поддерживает ее уровень. Внутриклеточный медиатор для ПЛ не известен.
Хорионический соматомаммолиберин (плацентарный лактоген) - не выполняет строго специфических функций, стимулирует лактацию и лютеотропную активность. Эффекты метаболизма такие же, как и у гормона роста.
Гликопротеиновые гормоны гипофиза и плаценты:
1. тиреотропный гормон (ТТГ);
2. лютеинизирующий гормон (ЛГ);
3. фолликулостимулирующий гормон (ФСГ);
4. хорионический гонадотропин (ХГ или ВХГ или ХГЧ).
Первые 3 гормона вырабатываются гипофизом, а ХГ – плацентой.
Эти 4 гормона структурно близки, они действуют через цАМФ. ФСГ, ЛГ, ХГ - гонадотропины, т.е. они действуют на половые железы.
ФСГ связывается с клетками-мишенями (у женщин - фолликулярные клетки яичников, у мужчин - клетки Сертоли семенников). Стимулирует рост фолликулов, подготавливает их к действию ЛГ. В мужском организме ФСГ индуцирует синтез андроген-связывающего белка, стимулирует рост семенников, семенных канальцев, сперматогенез.
ЛГ (лютропин). Клетки-мишени - клетки желтого тела у женщины и клетки Лейдена у мужчин. ЛГ стимулирует образование прогестерона у женщин и тестостерона (из холестерина) у мужчин. У женщин стимулирует овуляцию.
ХГЧ - хорионический гонадотропин человека - плацентарный гормон, синтезируется в плаценте и по строению близок к ЛГ. Его содержание возрастает в крови и в моче после имплантации (ранний признак беременности).
ТТГ. Клетки-мишени - клетки щитовидной железы. Увеличивает синтез трийодтиронина (Т3) и тироксина (Т4). Стимулирует включение йода и гидролиз тиреоглобулина с образованием трийодтиронина и тироксина. ТТГ увеличивает синтез белка в ЩЖ, синтез нуклеиновых кислот, что приводит к увеличению числа и размеров тиреоидных клеток. Образование ТТГ зависит от уровня периферических гормонов - Т3 и Т4. Если их концентрация увеличивается, то синтез ТТГ снижается.
Семейство проопиомеланокортина (ПОМК)
ПОМК - это молекула-предшественник, состоит из 285 АК-остатков. Этот белок гидролизуется до пептидов, которые и имеют гормональный эффект.
Выделяют 3 группы пептидов:
1. АКТГ (адренокортикотропный гормон) Из него могут образовываться a-МСГ (меланоцитстимулирующий гормон) и кортикотропиноподобный пептид;
2. b-липотропин - из него образуются a-липотропин, b-МСГ, b-эндорфины (из него - a-эндорфины, g-эндорфины);
3. N-концевой пептид, из него образуется g-МСГ.
АКТГ - полипептид, состоит из 39 АК. Клетки-мишени - клетки коры надпочечников. Повышает синтез стероидов коры надпочечников за счет превращения холестерола в прегненалон. Его содержание значительно возрастает - это предшественник кортикостероидов. Повышает синтез белков и РНК в коре надпочечников. Внутриклеточный посредник - цАМФ.
АКТГ стимулирует синтез аденилатциклазы в надпочечниках и жировой ткани, следовательно, интенсивность липолиза повышается. При избытке АКТГ - синдром Кушинга - повышается пигментация кожи, наблюдается отрицательный азотистый баланс, отрицательный баланс К и Р, но задерживается в организме натрий, что приводит к развитию отеков, повышению давления. Гиперсекреция АКТГ повышает уровень глюкозы в крови и может развиться стероидный диабет, повышается уровень свободных жирных кислот в крови, так как повышается липолиз - это характерно для стрессовых ситуаций.
b-липотропин - стимулирует липолиз.
b-эндорфины - в гипофизе они ацилированны и неактивны, а в ЦНС они свободны и проявляют активность. Они выполняют нейромедиаторную функцию и нейрорегуляторную. Эндорфины имеют те же рецепторы, что и морфины. Эндорфины регулируют чувство удовольствия (счастья). Применение различных опиатов приводит к увеличению числа рецепторов и возникает зависимость от внешних веществ (если количество рецепторов снижается).
МСГ - стимулирует меланогенез, т.е. образование меланоцитов и меланина.
Гормоны задней доли гипофиза (нейрогипофиза):
- вазопрессин (антидиуретический гормон);
- окситоцин.
Синтезируются в гипоталамусе, а выделяются из задней доли гипофиза.
Вазопрессин - по структуре пептид, состоящий из 9 АК.
Эти гормоны влияют на сокращения гладкой мышечной ткани, но клетки-мишени различны. Для вазопрессина это клетки сосудов и дистальных извитых канальцев и извитых трубочек почек. В связи с этим вазопрессин повышает обратное всасывание воды (реабсорбцию) в почках, т.е. диурез снижается и повышается сокращение гладкомышечных клеток сосудов. Он действует через вторичный посредник - цАМФ. При нарушении секреции вазопрессина может развиться несахарный диабет (несахарное мочеизнурение - нарушается реабсорбция и объем воды повышается - полиурия, но в моче нет глюкозы).
Окситоцин действует на гладкомышечную ткань матки и молочных желез. Посредник неизвестен. Используется для стимуляции родовой деятельности, оптимизирует секрецию молока.
Гормоны щитовидной железыВ щитовидной железе образуются 3 гормона, 2 из которых близки по действию и образуются из тирозина:
1. 3,5,3’-трийодтиронин (Т3);
2. 3,5,3’,5’–тетрайодтиронин (Т4);
3. тиреокальцитонин.
Т3 и Т4 образуются из АК – тирозина.
Образование тиреотропных гормонов происходит на белке тиреоглобулине. Сначала молекулы тирозина включаются в белковую структуру.

Особенности образования тиреоидных гормонов:
- они связаны со структурой тиреоглобулина в ЩЖ;
- для синтеза необходим йод.
ЩЖ способна концентрировать йод в виде йодидов, при этом происходит:
1. окисление йодидов с помощью тиреопероксидазы;
2. йодирование тирозина;
3. конденсация (соединение) йод-тирозина.
Тиреоглобулин, который содержит Т3 и Т4, разрушается с образованием этих гормонов под влиянием тиреотропного гормона (тиреоглобулин«(над стрелкой ТТГ, под - KI) Т3+Т4. KI тормозит распад тиреоглобулина и поэтому применяется для лечения гипертериоза.
В кровь поступают свободные тиронины, но в крови они транспортируются с тироксин-связывающим глобулином (ТСГ). Биологически более активны свободные тиронины, но их немного.
Механизм действия тиреоидных гормонов.
Тиреоидные гормоны проникают через плазматическую мембрану и связываются с ядерными рецепторами (рецепторы есть в ядре и цитоплазме). Сродство Т3 в 10 раз больше, чем Т4. Эти гормоны (Т3, Т4) индуцируют синтез белков, активируют транскрипцию мРНК.
Влияние на обмен веществ:
1. увеличивают поглощения кислорода почти во всех тканях, кроме мозга и половых желез;
2. повышают активность Na+,K+-АТФ-азы, но если активность повышается, концентрация АТФ снижается, следовательно усиливаются окислительные процессы;
3. снижают отношение кислорода к АТФ.
Тиреоидные гормоны повышают синтез белка, при их нормальной концентрации наблюдается положительный азотистый баланс, но при очень высокой концентрации этих гормонов проявляется обратный эффект.
Нарушения секреции.
Гипофункция – гипотиреоз - у детей может привести к нарушению умственного и физического развития, кретинизм. У взрослых - микседема (слизистый отек). Развитие эндемичного зоба связано с недостатком йодидов в воде и пище. Железистая ткань не развивается, а вместо нее образуется соединительная ткань, что приводит к увеличению массы ЩЖ.
Гиперсекреция тиреоидных гормонов – гипертиреоз – проявляется в виде тиреотоксикоза - увеличение ЩЖ, но за счет железистой ткани. При этом повышаются окислительные процессы в организме. Наблюдается отрицательный азотистый баланс, потеря массы тела, повышается возбудимость, пучеглазие, дрожание конечностей.
Кальцитонин образуется в клетках ЩЖ. Это пептид, состоящий из 32 АК. Он снижают уровень Са2+ в крови, но механизм действия недостаточно изучен.
Гормоны паращитовидных железПарат-гормон, состоит из 84 АК, регулирует уровень Са2+, стимулирует выход кальция (и фосфора) из костей в кровь; Повышают реабсорбцию кальция в почках, но стимулируется выход фосфора; Стимулирует образование кальцитриола из витамина Д3 в почках; Повышает всасывание кальция в кишечнике. Действует через цАМФ.
Гормоны поджелудочной железы (ПЖ)
ПЖ обладает и экзокринной и эндокринной функцией. В островках Лангерганса находятся клетки А, B, D, F. Инсулин - 70% - B-клетки; глюкагон - 25%- А-клетки; соматостатин - около 5% - D-клетки; панкреатический полипептид (PP)– следы - F- клетки.
Инсулин – это полипептид, состоящий из 51 АК. Состоит из двух полипептидных цепей: А и B-цепи. Эти цепи соединяются дисульфидными мостиками. [рис. 2-х цепей. А-цепь на левом (N-конце) завершается ГЛИ, на правом (С-конце) – АСП, 21 АК; В-цепь слева ФЕН, справа ТРЕ, 30 АК. 2 дисульфидных мостика]
Инсулин синтезируется в виде проинсулина, который состоит из 84 АК - синтезируется в виде 1 полипептидной цепи. При превращении проинсулина в инсулин отщепляется С-цепь из 33 АК (проинсулин®инсулин+С-пептид).
Клетки-мишени - мышечная ткань, печень, жировая ткань. Рецепторы к инсулину находятся на поверхности клеток. Рецепторы могут подвергаться интернализации, т.е. проникать внутрь мембраны комплексов «инсулин- рецептор», что приводит к разрушению путем эндоцитоза. Это объясняется снижением чувствительности клеток организма к инсулину при ожирении, т.к. количество рецепторов снижается и возникает устойчивость к инсулину. Внутриклеточный медиатор к инсулину не известен. Инсулиновые рецепторы являются ферментами и повышают активность фосфодиэстеразы и снижают уровень цАМФ.
Влияние инсулина на метаболизм:
1. углеводный обмен:
- снижение уровня глюкозы;
- повышение транспорта глюкозы через мембрану мышц и жировую ткань, без повышения специфических переносчиков глюкозы в печень.
- в печени активируется глюкокиназа, т.е. глюкоза®(над стрелкой инсулин) глюкозо-6-фосфат;
- инсулин повышает интенсивность утилизации глюкозы - гликолиз;
- активирует гликогенсинтазу, повышая синтез гликогена;
- снижает активность глюкозо-6-фосфатазы, т.е. глюкоза не выходит из печени;
- инсулин ингибирует глюконеогенез.
Т.е. инсулин стимулирует усвоение глюкозы организмом. 30-40% глюкозы превращается в жиры, 50% идет на гликолиз, а 10% превращается в гликоген.
2. липидный обмен:
- стимулирует липогенез;
- ингибирует липолиз.
3. белковый обмен:
- повышает синтез белка;
- ингибирует распад белка;
- стимулирует транспорт АК в клетки;
- повышает транскрипцию мРНК;
- повышает пролиферацию клеток, усиливая факторы роста фибробластов, тромбоцитарного фактора роста и фактора роста эпидермиса.
Дефицит инсулина – гипосекреция - развитие сахарного диабета. Есть 2 вида сахарного диабета:
1. недостаток инсулина - инсулинзависимый сахарный диабет - 10% (ИНЗСХ) - нарушение секреции инсулина в следствие генетических нарушений или поражение поджелудочной железы;
2. устойчивость к действию инсулина - инсулиннезависимиый сахарный диабет - 90% (ИнНСД)- снижение количества рецепторов к инсулину за счет интернализации рецепторов. Наблюдается при ожирении и повышенном потреблении сахара.
Клинические проявления одинаковы:
- гипергликемия - в клетках углеводное голодание, распад белков и жиров, стимуляция глюконеогенеза;
- глюкозурия – содержание глюкозы в крови больше 10 ммоль/л;
- полиурия;
- полидипсия - избыточное потребление воды;
- полифагия - повышенный голод;
- кетоз;
- ацидоз (кетоацидоз);
- диабетическая кома.
Глюкагон – это полипептид, состоящий из 29 АК. Синтезируется в А-клетках поджелудочной железы. Клетки-мишени - печень. Рецепторы к глюкагону находятся на поверхности клетки, посредник - цАМФ. Глюкагон повышает уровень цАМФ, следовательно увеличивается протеинкиназа® увеличивается количество фосфорилазы-В® увеличивается количество фосфорилазы-А, которая действует на превращение гликогена в глюкозу.
Эти процессы активизируются в печени. Глюкагон повышает глюконеогенез, липолиз.
Панкреатический полипептид - функции неизвестны.
Инсулиноподобные факторы роста (ИФР) - не образуются в ПЖ, но близки по строению и функции к инсулину и влияют на рост и пролиферацию клеток. ИФР больше действуют на синтез белка и пролиферацию.
Соматостатин - образуется в гипоталамусе и в D-клетках ПЖ, а также в ЖКТ. Подавляет секрецию других гормонов ПЖ, т.е. обладает паракринным эффектом. Снижает секрецию гастрина в желудке, опорожнение желудка, снижает всасывание углеводов.
Гормоны коры и мозгового вещества надпочечников
Гормоны мозгового слоя - адреналин и норадреналин - производные тирозина. [схема образования: тирозин ® (оксигеназа, 1/2О2)диоксифенилаланин ® (декарбоксилаза, -СО2)дофамин ® (+О2)норадреналин ® (+СН3)адреналин].

Органы-мишени - печень, скелетные мышцы, сердечная мышца, слюнные железы, матка.
Механизм: через аденилатциклазную систему. Увеличивается цАМФ ® увеличение протеинкиназы ® увеличение фосфорилирования. Фосфорилазы действуют на превращение гликогена в глюкозу.
Адреналин действует и в печени, и в мышцах, вследствие чего повышается содержание глюкозы в крови, повышается содержания молочной кислоты. Адреналин повышает потребление кислорода, увеличивает липолиз, что приводит к росту количества свободных жирных кислот в крови. Повышает кровяное давление, частоту сердечных сокращений. Адреналин действует на a1, a2, b1 и b2-адренорецепторы. Если гормоны связаны с b-рецепторами, то происходит активация цАМФ, если с a-рецепторами - ингибирование цАМФ.
Гормоны коркового вещества надпочечников (кортикостероиды).
Известно более 30 гормонов-стероидов, т.е. производные циклопентанпергидрофенантрена:
1. глюкокортикоиды - оказывают влияние на углеводный обмен;
2. минералокортикоиды - на минеральный обмен;
3. половые гормоны.
Глюкокортикоиды: кортикостерон, кортизол (самый активный в организме человека), кортизон.

Клетки-мишени для глюкокортикоидов - печень, почки, лимфоидная ткань, соединительная ткань, мышцы.
Рецепторы находятся в цитозоле, проходят через мембрану и действуют на ген. Гормон ® ген ® белок.
Влияние на обмен веществ:
1. активация глюконеогенеза;
2. повышение уровня глюкозы в крови;
3. повышение синтеза гликогена в печени;
4. стимулируют липолиз в области конечностей и липогенез в области туловища и лица;
5. повышение окисления жирных кислот;
6. повышение образования кетоновых тел;
7. в печени увеличивается синтез белка, в мышцах, лимфоидной, соединительной тканях увеличивается распад белка;
8. противовоспалительное действие, вызывают инволюцию лимфоидной ткани;
9 антиаллергический эффект, подавляют образование антител;
10. подавление синтеза белка в соединительной ткани, задержка образования рубцов и спаек.
Использование глюкокортикоидов в клинике - противовоспалительные, при трансплантации органов для снижения образования рубцов и спаек.
Глюкокортикоиды влияют и на минеральный обмен, но в меньшей степени.
Минералокортикоиды:
- диоксикортикостерон;
- альдостерон.

Клетки-мишени - дистальные канальцы почек. Рецепторы находятся внутри клеток – цитозольные рецепторы. Влияют на синтез белков, транспорт натрия через мембраны, т.е. повышают реабсорбцию натрия и хлоридов из первичной мочи, задерживая натрий в организме. Влияют на углеводный обмен, как и гипоталамус, но в меньшей степени.
Гипофункция коры надпочечников - болезнь Адиссона (бронзовая болезнь). Снижается устойчивость организма к стрессам, гипогликемия, потеря натрия и накопление калия, гипотония, мышечная слабость, утомляемость, повышение пигментации кожи, возможна гибель из-за нарушения водно-солевого обмена.
Гиперфункция - синдром Иценко-Кушинга.
Гормоны половых железГонады - яйчники и семенники - железы смешанного типа. По химической природе их гормоны - стероиды. Андрогены (мужские) синтезируются в семенниках и коре надпочечников. Эстрогены (женские) - в яичниках и коре надпочечников. Синтезируются из холестерола, который образуется из Ац-КоА. Ац-КоА® холестерол® прегненолон ®прогестерон ®кортикостероиды. Из прогестерона также образуются андрогены (тестостерон) , а из них - эстрогены.
Эстрогены: эстрадиол, эстрон (образуется из эстрадиола в плаценте), эстриол, прогестерон.

Андрогены: тестостерон, андростерон (образуется из тестостерона).

Органы-мишени для эстрадиолов - матка и молочные железы, для андрогенов - простата, семенные пузырьки, мышцы.
Механизм действия – цитозольный - гормоны соединяются с цитозольными рецепторами и действуют на ген.
Основной эффект - синтез специальных белков репродуктивной системы.
Влияние на обмен веществ.
Эстрогены:
1. синтезируют специфические белки;
2. обуславливают положительный азотистый баланс;
3. активируют пентозный цикл;
4. препятствуют накоплению липидов в жировой ткани и печени.
Эстрогены имеют цикличность секреции: в фолликулярной фазе синтезируются эстрогены, в лютеиновой фазе - прогестерон.
Андрогены:
1. синтезируют специальные белки половой сферы;
2. стимулируют синтез мышечных белков;
3. стимулируют синтез белков печени и почек;
4. ускоряют развитие костной ткани;
5. ускоряют аэробное окисление;
6. обладают анаболическим эффектом;
7. влияют на развитие половых органов, вторичных половых признаков, половое влечение.
Витамины
К концу XIX века сложились представления, что основными компонентами пищи являются белки, жиры, углеводы и минеральные вещества. Но ряд наблюдений указывал на недостаточность только этих групп питательных веществ. (Напр., Лунин, 1880: при кормлении одной группы мышей естественными продуктами, а другой – белками, жирами, углеводами, полученными из этих же продуктов, гибель мышей во второй группе была значительно выше.)
В 1911 г. польский ученый Функ выделил в рисовых отрубях соединение, содержавшее аминогруппу, которое было необходимо для нормальной жизнедеятельности. Он назвал его витамином (от лат. vitae amine – амин жизни).
Витамины – это необходимые для нормальной жизнедеятельности низкомолекулярные органические соединения, синтез которых в организме ограничен или отсутствует.
Отличительные признаки витаминов:
1) витамины не выполняют пластических функций;
2) витамины не играют энергетической роли (т.е. не используются как источник энергии);
3) витамины не синтезируются в организме или синтезируются в недостаточных количествах;
4) дефицит витаминов вызывает специфические нарушения обмена веществ с характерными клиническими проявлениями;
5) выполняют специфические функции, которые невозможно заменить другими органическими соединениями;
6) витамины необходимы в миллиграммах или микрограммах в сутки (!).
Классификация витаминовПринята классификация по растворимости:
1) жирорастворимые (A, D, E, K);
2) водорастворимые (все остальные: В_, С, Р, Н);
3) витаминоподобные вещества – синтезируются в организме, но в недостаточном количестве (парааминобензойная к-та, коэнзим Q, холин, инозит, пангамовая к-та).
Номенклатура витаминов представлена химическим названием и буквенным обозначением.
Напр., витамин А – ретинол, витамин РР – никотинамид или никотиновая кислота, витамин В6 – пиридоксин, пиридоксаль или пиридоксамин.
Также существует клиническое название витамина, которое состоит из названия патологического проявления недостатка витамина с приставкой анти- .
(напр., витамин А – антиксерофтальмический витамин, витамин D – антирахитический витамин).
Жирорастворимые витамины могут откладываться в печени в виде резервов (A, D, K), витамин Е может накапливаться в жировой ткани. Так как эти витамины нерастворимы в воде, то они не могут свободно проникать в кровь и выводиться с мочой. При избыточном поступлении этих витаминов могут развиваться токсические эффекты (в основном, витамина А и D).
Водорастворимые витамины выводятся с мочой, поэтому их избытка не наблюдается. Возможен небольшой резерв фолиевой к-ты и витамина В12 в печени. Токсические эффекты не наблюдаются.
Роль витаминов в обмене веществ1.(!) витамины – предшественники коферментов и простетических групп ферментов.
Напр., В1 – тиамин – входит в состав кофермента декарбоксилаз кетокислот в виде ТПФ (ТДФ), В2 – рибофлавин – входит в состав коферментов дегидрогеназ в форме ФМН и ФАД, РР – никотинамид – входит в состав НАД и НАДФ – коферментов дегидрогеназ;
2. витамины – предшественники гормонов (витамин D3 – предшественник кальцитриола);
3. витамины могут использоваться для синтеза медиаторов (витамин С – для синтеза норадреналина и серотонина);
4. витамины участвуют в образовании гормонов (витамин С – в образовании кортикостероидов);
5. витамины необходимы для синтеза незаменимых АК (витамин В12 – метионин).
Понятие о гиповитаминозах, авитаминозах и гипервитаминозахГиповитаминоз – патологическое состояние, связанное с недостатком витамина в организме.
Авитаминоз – патологическое состояние, вызванное отсутствием витамина в организме.
Полигиповитаминоз – патологическое состояние, связанное с недостатком нескольких витаминов в организме.
Гипервитаминоз – патологическое состояние, связанное с избытком каких-либо витаминов в организме (обычно витаминов А или D).
Чаще присутствует недостаточная витаминная обеспеченность, т.е. отсутствуют характерные клинические проявления, а лишь снижена работоспособность, повышена сонливость, увеличивается частота простудных заболеваний и их тяжесть, появляются заболевания сердечно-сосудистой системы и т.д.
Причины гиповитаминозов1. Первичные: недостаток витамина в пище.
2. Вторичные:
а) снижение аппетита;
б) повышенный расход витаминов;
в) нарушения всасывания и утилизации, напр., энтероколиты, желчно-каменная болезнь;
г) химиотерапия (антибиотики, аспирин – недостаток фолиевой к-ты);
д) хирургические вмешательства;
е) физиотерапия и др.
3. Врожденные: отсутствие ферментов, катализирующих превращение провитамина в витамин (напр., каротин не превращается в ретинол – витамин А).
Причины недостаточной витаминной обеспеченности в современных условиях
Чешские исследователи выяснили, что при оптимальной диете человек не получает достаточное количество витаминов. Причины:
1) снижение энерготрат с 3500-4000 ккал/сут до 2000-2500 ккал/сут за сто лет, из-за чего снижение количества пищи и, следовательно, уменьшение количества витаминов;
2) использование бедных витаминами продуктов (рафинированных и высококалорийных: сахар, белый хлеб, спирт);
3) использование рафинированных продуктов;
4) недостаточное потребление овощей и фруктов;
5) отсутствие централизованной системы витаминизации.
Т.о., рацион современного человека достаточен по калорийности, но не удовлетворяет его потребностей в витаминах.
Жирорастворимые витаминыВитамин А
Витамеры: А1 – ретинол и А2 – ретиналь.
Клиническое название: антиксерофтальмический витамин.
По химической природе: циклический непредельный одноатомный спирт на основе кольца b-ионона.

Может разрушаться кислородом, т.е. является антиоксидантом.
Роль витамина А в метаболизме:
1. Участвует в росте и дифференцировке клеток эмбриона, развивающегося организма. Участвует в делении и дифференцировке быстро пролиферирующих тканей (хрящевые, костные, эпителиальные ткани), т.к. витамин А может инициировать репликацию и участвует в образовании хондроитинсульфата.
2. Участвует в фотохимическом процессе зрения. В состав зрительного пигмента родопсина входит 11-цис-ретиналь, который при освещении переходит в 11-транс-ретиналь, активирующий фосфодиэстеразу, которая расщепляет цГМФ, в результате чего ионные каналы мембраны закрываются, возникает гиперполяризация мембраны и генерируется нервный импульс. При этом родопсин разлагается на белок опсин и 11-транс-ретиналь. В темноте наблюдается регенерация родопсина: транс-ретиналь® (алкоголь-ДГ, НАД·Н2 → НАД) транс-ретинол® (изомераза) цис-ретинол ®(алкоголь-ДГ, НАД·Н2 → НАД) цис-ретиналь® (+опсин) родопсин
Гиповитаминоз А проявляется нарушением темновой адаптации. Если наблюдается в растущем организме, то имеет место задержка роста. Гиповитаминоз А может перерасти в авитаминоз А, проявляющийся гемеролопией ("ночной слепотой"). При недостатке витамина А нарушается эпителизация, наблюдается избыточное ороговение эпителия (сухость кожи, сухость роговицы глаз – ксерофтальмия). Ксерофтальмия может привести к развитию микрофлоры, кератомаляции (размягчению роговицы), затем к её помутнению и амблеопии (слепоте).
Причины гипо- и авитаминозов А:
- недостаток в пище
- нарушение всасывания в кишечнике
- заболевания печени, при которых провитамин (кератин) не превращается в витамин
- повышенная потребность в витамине А – у растущего организма, беременных, ночных водителей)
Суточная потребность витамина А 1,0 - 2,5 мг, а провитамина А (каротина) 2,0 – 4,0 мг.
Каротин (провитамин А) – димер витамина А. Содержится в растительных продуктах. (Различают альфа-, бета- и гамма-каротин.)
Источники витамина А:
- животные жиры (рыбий жир, печень, яичный желток, сливочное масло);
- каротин растительного происхождения (морковь, свекла, томаты, зеленый горошек).
Витамин D
Антирахитический витамин. Существуют два витамера:
D2 – эргокальциферол и D3 – холекальциферол.
Витамин D2 содержится в грибах. Витамин D3 синтезируется в организме под действием УФО (ультрафиолетового облучения):
7-дегидрохолестерол → холекальциферол (D3)

Роль витамина D в обмене веществ:
Витамин D3 (холекальциферол) подвергается в организме превращению. Он поступает в печень, где под действием 25-гидроксилазы превращается в 25-гидроксихолекальциферол, затем в почках под действием ПТГ и 1-гидроксилазы – в 1,25-дигидроксихолекальциферол (гормон кальцитриол)

кальцитриол
Функции кальцитриола
1. регулирует всасывание кальция и фосфора в кишечнике путем активации синтеза в энтероцитах кальций-связывающего белка (кальбиндина D);
2. в костях способствует минерализации ткани, поддержанию нормальной концентрации кальция и фосфора в межклеточном пространстве.
(повышает активность щелочной фосфатазы, повышает концентрацию кальций-связывающего белка и остеокальцина)
Гиповитаминоз D приводит к снижению уровня кальция в крови, снижению кальция и фосфора в межклеточном пространстве, нарушению минерализации костной ткани.
Авитаминоз D – рахит. Различают рахит I типа (при недостатке витамина D) и рахит II типа (при недостатке рецепторов). Проявляется рахит деформациями скелета ("рахитические четки", Х-образные или О-образные голени, килевидная грудная клетка).
Недостаток витамина D у взрослых сопровождается остеомаляцией, а в крови увеличивается активность щелочной фосфатазы.
Причины гиповитаминозов D:
- недостаток витамина D в пище
- недостаток УФ (солнечного) облучения
- недостаток парат-гормона
Суточная потребность вит. D 0,012-0,025 мг.
Источники витамина D: яичный желток, рыбий жир, сливочное масло (лучше летнее), печень, молоко.
Витамин ЕУстар.: антистерильный витамин, антиоксидантный энзим.
В химическом плане это альфа-, бета-, гамма- и дельта-токоферолы, но преобладающим является альфа-токоферол.
Витамин Е устойчив к нагреванию.
Роль витамина Е в обмене веществ1. регулирует интенсивность свободно-радикальных реакций. Препятствует перекисному окислению липидов биомембран;
2. повышает активность витамина А.
Гиповитаминоз Е специфической картины не имеет. Наиболее специфичны гемолитическая анемия недоношенных и патологии мембран.
Суточная потребность витамина Е 20-25 мг.
Источники витамина Е: растительные масла (!), а также печень, желток яиц, проросшие зерна; масло облепихи.
Витамин КАнтигеморрагический витамин.
Витамеры: К1 – филлохинон и К2 – менахинон.
Роль витамина К в обмене веществЭто кофактор карбоксилирования глутаминовой кислоты (ГЛУ) в белке крови протромбине для его превращения в тромбин.
протромбин → тромбин [карбоксилирование гамма-углеродного звена остатка глутамата]
Антагонист витамина К – варфарин (крысиный яд), он близок по структуре к витамину К, является антикоагулянтом.
Витамин К поступает в организм с зелеными растениями (шпинат, крапива), жирами, а также синтезируется микрофлорой кишечника.
Гиповитаминоз К проявляется геморрагиями.
Авитаминоз К чаще наблюдается при нарушении всасывания его в кишечнике.
{водорастворимая форма витамина К - викасол}
Водорастворимые витамины Витамин САскорбиновая кислота, антискорбутный витамин (скорбут = цинга).
Является лактоном. Легко окисляется:
О=С─┐ О=С─┐
| │ | │
НО-С │ -2Н О=С │
║ О ◄═══► | О
НО-С │ +2Н О=С │
| │ | │
НС─┘ НС─┘
| |
НО-СН НО-СН
| |
Н2С-ОН Н2С-ОН
аскорбат дегидроаскорбат
Енольные гидроксилы неустойчивы, особенно в присутствии кислорода. В кислой среде витамин С сохраняется лучше.
Роль витамина С в обмене веществ1. участвует в реакциях гидроксилирования (ЛИЗ→ОЛИ, ПРО→ОПР), что требуется для "сшивок" молекул коллагена в соединительной ткани;
2. участвует в триптофана и диоксиметила с образованием нейромедиаторов (норадреналин и др.);
3. необходим для синтеза кортикостероидов;
4. необходим для образования нейромедиаторов.
Гиповитаминоз С проявляется поражением соединительной ткани – повышенная хрупкость сосудов, пипехии (точечные кровоизлияния на коже), кровоточивость десен.
Авитаминоз С – цинга (скорбут): кровоизлияния во внутренние органы, более выраженные повреждения соединительной ткани.
Суточная потребность витамина С 100-150 мг. Она возрастает при инфекционных заболеваниях, стрессовых состояниях, лактации, беременности – до 300 мг/сут.
Источники витамина С: овощи, фрукты, зеленые растения; главные – черная смородина, шиповник, грецкий орех, цитрусовые. В России основным источником его является картофель (10 мг/100 г) и квашенная капуста.
Причины гиповитаминоза С:
1. витамин С неустойчив;
2. он необходим в относительно больших количествах;
3. в организме отсутствуют запасы витамина С.
Витамин РВитамин проницаемости.
Витамеры: рутин, катехины, цитрин.
Это витамин-спутник витамина С.
Гиповитаминоз Р: поражение сосудов, нарушение их проницаемости.
Роль в обмене веществ
Витамин Р тормозит активность гиалуронидазы, чем сохраняет соединительную ткань. А также участвует в окислительно-восстановительных реакциях.
Потребность в витамине Р не установлена.
Витамин В1
Тиамин, антиневритный витамин.

Тиамин устойчив в кислой среде (до 140ºС), а в щелочной среде быстро разрушается.
Роль витамина В1 в обмене веществ
1. из него образуется ТПФ (тиаминпирофосфат) – кофермента декарбоксилаз кетокислот (пируват-ДК-комплекс, альфа-КГ-ДК) и транскетолазы;
2. участвует в передаче нервного импульса;
3. является коферментом транскетолазы.
Гиповитаминоз В1: накопление ПВК и альфа-КГ в крови из-за нарушения их превращений, поражение нервной ткани из-за недостатка глюкозы.
Авитаминоз В1 – болезнь бери-бери: полиневриты, отеки, сердечно-сосудистая недостаточность (иногда до некрозов), нарушения секреции и моторики ЖКТ (атония кишечника). Чаще развивается при хроническом алкоголизме, когда витамин В1 не всасывается.
Суточная потребность витамина В1 2-3 мг. Потребность возрастает при углеводной пище (0,5 мг витамина на каждые 1000 ккал).
Источники тиамина: дрожжи, хлеб грубого помола, каши, крупы (овсяная, гречневая, фасоль).
Витамин В2
Рибофлавин

Устойчив в кислой среде, но разрушается в нейтральной и щелочной. Легко окисляется по двойной связи, что позволяет ему участвовать в о/в реакциях в виде коферментов (ФМН, ФАД):
- окисляет восстановленную форму НАД·Н (компонент НАД·Н-дегидрогеназы в дыхательной цепи),
- окисляет жирные к-ты, янтарную к-ту, аминокислоты.
Авитаминоз В2: поражение эпителия слизистых, кожи, глаз; сухость слизистых губ, полости рта, трещины губ; дерматиты, сухость конъюнктивы, переходящая в конъюнктивиты, кератиты, васкуляризация глазных яблок.
Суточная потребность 2-4 мг.
Источники: дрожжи, печень; хлеб грубого помола, соя, яйца, молоко.
Витамин РРАнтипеллагрический витамин.
Витамеры: никотиновая к-та, никотинамид, ниацин.


Устойчив при кипячении.
Роль витамина РР в обмене веществ
Используется для синтеза НАД и НАДФ – коферментов дегидрогеназ.
Гиповитаминоз РР: усталость, слабость. Более выраженный – пеллагра: диарея, дерматиты, деменция (слабоумие).
Суточная потребность витамина РР 20-25 мг.
Источники: дрожжи, печень, грибы, соя, бобы, мясо, мука пшеничная грубого помола. Может синтезироваться в организме из аминокислоты триптофана при участии витамина В6.
Поэтому гиповитаминоз РР бывает при белковом голодании и при гиповитаминозе В6.
Витамин В6
Антидерматитный витамин.
Пиридоксин → пиридоксаль → пиридоксамин [нарисовать формулы]

(все три эти соединения обладают витаминным действием)
Роль витамина В6 в обмене веществ
1. он необходим для образования ПФ (пиридоксальфосфата) – кофермента амино-ТФ, декарбоксилаз АК, дезаминаз АК;
2. необходим для превращения триптофана в витамин РР;
3. нужен для превращения дельта-аминолевулиновой кислоты в гем.
{Т.е., вит.В6 нужен для обмена аминокислот}
Гиповитаминоз В6: анемия из-за нарушения обмена АК, плюс дерматиты, стоматиты, глосситы, конъюнктивиты.
Суточная потребность в витамине В6 2-3 мг.
Источники: печень, дрожжи, хлеб грубого помола, горох. Также он синтезируется микрофлорой кишечника.
Гиповитаминоз возможен при длительном употреблении антибиотиков, особенно, противотуберкулезных препаратов.
Витамин В9, В10, ВС (фолиевая кислота)
Фолиевая кислота, фолацин. Антианемический витамин.
Роль фолиевой кислоты в обмене веществ
Участвует в транспорте одноуглеродных фрагментов (-СООН, -СН3, СН=О): превращение урацила в тимин, этаноламина в холин; синтез АК метионина, серина, пуриновых оснований.
{Т.е., фолиевая кислота нужна для обмена белков и нуклеиновых кислот}
Гиповитаминоз фолиевой кислоты: анемия, затем диарея.
Суточная потребность фолиевой к-ты 0,2 мг.
Источники: зеленые растения (шпинат, капуста), фасоль, печень, молоко, яйца; плюс синтезируется микрофлорой кишечника.
Причины недостатка фолиевой к-ты:
- использование антибиотиков;
- заболевания ЖКТ (кишечника);
- недостаточность белкового питания.
Особенно недостаток фолиевой к-ты проявляется при беременности (у 58% беременных), им нужно до 0,6 мг/сут. Считается, что 98% недоношенных и имбицильных детей обусловлено дефицитом фолиевой кислоты.
Витамин В12
Кобаламин. Антианемический витамин.
Имеет красный цвет. На свету разлагается.
Роль кобаламина в обмене веществ
- транспорт метильных групп;
- участвует в транспорте водорода;
- превращение рибозы в дезоксирибозу;
- участвует в синтезе метионина.
{Т.е., кобаламин нужен для обмена белков и нуклеиновых кислот.}
Недостаток кобаламина: анемия, поражение нервной системы, снижение кислотности желудочного сока.
Суточная потребность в кобаламине 0,003 мг.
Источники: печень, рыба, яйца, молоко, + синтезируется микрофлорой. (в дрожжах – нет)
Недостаток кобаламина может возникать при заболеваниях желудка, в котором вырабатывается внутренний фактор Касла (транскоррин), необходимый для всасывания витамина В12.
Витамин В3
Пантотеновая кислота. [рис. формулы НОСН2-С((СН3)2)-СН(ОН)-СО-NH-СН2-СН2-СООН]
Состоит из масляной кислоты с b-аланином.
Роль в обмене веществ
Участвует в образовании КоА, который в свою очередь участвует:
1. синтезе и распаде жирных кислот;
2. транспорте ацильных остатков (пр., АцКоА)
3. в реакциях окислительного декарбоксилирования пирувата и a-кетоглутаровой кислоты.
{Т.е. витамин В3 необходим для обмена Б, Ж, У}
Авитаминоз – представлен разными формами; характерны дерматиты, поражения слизистой оболочки внутренних органов, поражение ЖВС (надпочечники, тимус), поражение нервной ткани. Авитаминозы практически не встречаются.
Суточная потребность 10 мг.
Этот витамин широко распространен, содержится в печени, яйцах. овсяных хлопьях, дрожжах. Также синтезируется микрофлорой кишечника.
Витамин Н (биотин)Антисеборейный витамин. Структура: тиофен, соединеный с мочевиной+ боковая цепь (валериановая кислота).
Устойчив при нагревании.
Роль биотина в обмене веществ
Биотин – кофермент карбоксилирования (превращение ацетил-КоА в малонил-КоА в синтезе ВЖК) и кофермент транскарбоксилирования в синтезе пуриновых оснований.
Авитаминоз Н – себорея: покраснение и шелушение сальной кожи на волосистой части головы.
Суточная потребность витамина Н 0,010 мг.
Источники: печень, яйца, молоко.
Авитаминоз бывает при употреблении сырых яиц, содержащих авидин (антивитамин Н).
Витаминоподобные вещества
Парааминобензойная кислота
Роль: входит в состав фолиевой кислоты, участвует в пигментации. При авитаминозах нарушение пигментации.
ХолинОтносится к витаминоподобным веществам.
НО-СН2-СН2-N(CH3)3
Роль в обмене веществ
1. холин входит в состав сложных липидов (фосфатидилхолин);
2. источник метильных групп при синтезе различных соединений.
В организме синтезируется из серина и метионина в необходимых количествах. Но метионин – незаменимая АК!
АНТИВИТАМИНЫ
Это вещества, которые нарушают использование витаминов.
Их 2 группы:
1. антивитамины, прямо воздействующие на снижение активности витаминов, например авидин на витамин А, аскорбатоксидаза на витамин С;
2. структурные аналоги витаминов, похожие на витамины, но оказывающие противоположное действие, например сульфаниламиды похожи по структуре на ПАБК. Такие вещества используют для гибели микроорганизмов, которые их не различают.
Биохимия печени
Роль печени в обмене веществ
Печень занимает центральное место в обмене веществ организма. Особенность анатомического расположения и связи с другими органами, позволяет печени участвовать во всех видах обмена веществ. Печень кровоснабжается печеночной артерией и воротной веной, которые в ней образуют обширную капиллярную сеть, следовательно, все вещества поступающие в организм вместе с пищей, после их всасывания в кровь распределяются печенью по всему организму. Через печень протекает около 1,2 литра крови в минуту. Площадь капиллярных синусоидов в печени взрослого человека 400-450 м2.
Важнейшие функции печени:
1. синтетическая - происходит превращение питательных веществ в такие формы, которые могут быть использованы другими тканями:
- белки плазмы крови (альбумины, глобулины, фибриноген, протромбины);
- группа белков участвующих в транспорте липидов (ЛПОНП, ЛПНП, ЛПВП);
- гормонов (транскортины);
- витаминов;
- микроэлементы (трансферины, церрулоплазмины – переносчики ионов меди)
В печени синтезируется глюкоза, гликоген, нейтральные жиры, ХС, ФЛ, ВЖК (заменимые), кетоновые тела.
В печени активно синтезируются из АК небелковые азотистые соединения: коферменты (HS-KoA), креатин, глутатион, никотиновая кислота, пуриновые и пиримидиновые основания, порфирины.
В печени происходит биосинтез мочевины, мочевой кислоты, активно протекают реакции обмена АК (переаминирование, окислительное дезаминирование, декарбоксилирование), что обеспечивает поддержание в организме баланса заменимых АК, биогенных аминов.
2. депонирующая. В печени может депонироваться гликоген, ТГ, жирорастворимые витамины (А, D, Е, К), некоторые водо-растворимые витамины (В1, В2, В12), микроэлементы.
3. пищеварительная - связана с образованием и секретированием желчи и желчных кислот.
4. выделительная (экскреторная) – связана с образованием специально жидкого экскрета - желчи. Вместе с желчью из клеток удаляются избытки ХС, желчные кислоты, продукты распада гема (желчные пигменты), инактивированные витамины и гормоны, обезвреженные токсические вещества, вода.
5. обезвреживающая функция. В печени обезвреживаются токсические метаболиты обмена веществ (аммиак, крезол, фенол, скатол, индол, билирубин). В печени обезвреживаются, поступающие из вне через ЖКТ, вдыхаемый воздух, кожу - спирты, карболовые кислоты, альдегиды.
Поступающие в организм извне чужеродные вещества, а так же соединения, образующиеся в организме в процессе метаболических реакций, которые не могут быть использованы для пластических целей или как источники энергии, называются ксенобиотиками. Эти соединения в подавляющем большинстве случаев должны подвергаться обезвреживанию, т.е. детоксикации. Главным органом, где происходят эти процессы, является печень.
Кроме ксенобиотиков, инактивации в печени и выведению из организма подвергаются и некоторые собственные метаболиты обмена (продукты распада гема, стероидные гормоны, катехоламины). Все эти соединения из организма выводятся в виде метаболитов, химического комплекса с биомолекулами (с глутатионом), выводятся металлы. Некоторые из них выводятся в неизмененном виде (ацетон), некоторые соединяются с макромолекулами и могут, накапливаясь в организме, оказывать токсическое действие, медленно выделяясь из организма. Так взаимодействуют с белками соли тяжелых металлов.
В большинстве случаев ксенобиотики выводятся из организма в виде парных безвредных соединений – конъюгатов. Если в молекулах, поступающих с током крови веществ в печень, имеются свободные функциональные группы –OH, –SH,–NH2, то эти вещества могут сразу же вступать в фазу конъюгации.
Если у ксенобиотиков таких функциональных групп нет, то они должны предварительно пройти стадию химической модификации. В процессе этих реакций, соединение может подвергаться окислению или восстановлению, циклизации, дециклизации, гидролизу и т.д. При этом, в структуре вещества происходит разрыв внутренних связей, либо вводят дополнительные функциональные группы или происходит высвобождение заблокированной функциональной группы. Таким образом, появление дополнительной функциональной группы делает ксенобиотики более гидрофильными и следовательно способны вступить в фазу конъюгации.
Важным этапом детоксикации поступивших в печень с током крови веществ является их окисление в микросомах ЭПС. Здесь в липидном слое биомембран локализуется система гидроксилированных ферментов, требующих наличие молекул кислорода, НАДФН2 (восстановленый), поступающих из пентозного цикла, флавопротеида (ФП) и цитохрома.
Гидроксилазы способствуют включению кислорода в молекулу субстрата. НАДФН2 является источником электронов и протонов. Введение ОН-группы в структуру ксенобиотика делает его более полярным и, следовательно, облегчает его последующее конъюгирование.
НАДФН2 – зависимая моноксигеназная цепь микросом состоит из:
1. ФП, коферментом которого является ФАД;
2. цитохрома Р-450.
ФП обладает НАДН2–дегидрогеназной активностью, причем ФАД акцентирует протоны и электроны. С ФП электроны транспортируются на цитохром Р-450, а протоны в окружающую среду.
Цитохром Р-450 является последним самоокисляющим звеном этой цепи. Он выполняет двойную роль:
1. активирует кислород путем переноса электронов;
2. использует активированный кислород для окисления вещества и образование воды.
В этом процессе 1 атом кислорода включается в окисляемое вещество, а другой связывает два протона из среды и входит в состав воды.
Гидроксилирование ксенобиотиков с участием микросомальной монооксигеназной системы
1. бензола:
[рис. бензол+ О2 +НАДФН2®(гидроксилаза, цитохром Р450) фенол + НАДФ+ Н2О]
2. индола:
[рис. индол+ О2 +НАДФН2®(гидроксилаза, цитохром Р450) индоксил + НАДФ+ Н2О]
Кроме микросом, химическое превращений ксенобиотиков может проходить в митохондриях, лизосомах, цитозоле гепатоцитов. Обезвреживание ксенобионтов в печени проходит с участием основных конъюгатов: глюкуроновой кислоты, серной кислоты, уксусной кислоты и глицина и соответствующих ферментов – ТФ.
Конъюгация происходит путем химического связывания ксенобиотиков с активными формами – УДФ-ГК (с глюкуроновой кислотой ), ФАФС, Ац-КоА и глицина.
При образовании конъюгатов значительно уменьшается количество токсических соединений, увеличивается водорастворимость, что облегчает выведение их из организма в составе мочи.
Конъюгацией с глюкуроновой кислотой подвергаются: ксенобиотики, стероиды, билирубин, витамины D2, D3.
С серной кислотой: ксенобиотики, стероиды, тирамины, токаферолы, продукты гниения белков.
С уксусной кислотой - все соединения, имеющие свободную аминогруппу.
С глицином - соединения имеющие СООН-группу.
Реакции конъюгации ксенобиотиков:
1. с глюкороновой кислотой (УДФ-ГК):
[рис. УДФ-ГК+ фенол®(ТФ) фенилглюкоуронид+ УДФ]
2. с серной кислотой:
[рис. индоксил+ НО3S-ФАФ (это ФАФС)®(ТФ) индоксилсульфат+ ФАФ]
3. с глицином:
[рис. бензойная кислота+ NH2-CH2-COOH (это глицин) ®(ТФ) гиппуровая кислота+ Н2О]


Пигментный обмен представляет собой совокупность сложных взаимопревращений окрашенных веществ тканей и жидкостей организма человека.
К пигментам относятся 4 группы веществ:
1. гем-содержащие пигменты, входят в состав ФАВ, т.н. дыхательных пигментов (гемоглобина, миоглобина, цитохромов, каталазы, пероксидазы);
2. представлены предшественниками гема – порфиринами;
3. представлены продуктами распада гема – желчными пигментами (билирубин желчи, уробилин мочи, стертобилин кала);
4. образуются из аминокислоты (ТИР) в пигментных клетках - меланоцитах. Это меланин. Он находится в коже, волосах, радужной оболочке глаз.
Т.о., большая часть пигментов человека – это гемохромагенные пигменты, которые образуются либо при синтезе, либо при распаде гема.
Биосинтез гемаБиосинтез гема идет в большинстве тканей, за исключением эритроцитов, которые не имеют митохондрий. В организме человека гем синтезируется из глицина и сукцинил-КоА, образованного в результате метаболических реакции ЦТК.
Для активации глицина необходим витамин В6. В процессе первой метаболической реакции синтеза гема образуется d-аминолевулиновая кислота (АЛК). Скорость синтеза гема контролируется ферментом – синтаза. Наиболее активно эта реакция идет в печени.
[рисуется в столбик. Первая реакция идет в митохондриях: глицин + сукцинил-КоА®(синтаза) аминолевулиновая кислота (АЛК). Следующие реакции в цитозоле: АЛК + АЛК® порфобилиноген (монопиррол) + 3 порфобилиногена ® уропорфобилиноген (или тетрапиррол) ® (над стрелкой: декарбоксилаза; под: -СО2) копропорфобилиноген. Далее реакции до образования гема идут опять в митохондриях: копропорфобилиноген ®(над стрелкой: декарбоксилаза; под: -СО2) протопорфобилиноген ® (над стрелкой оксидаза; под: +О2) протопорфирин ® (над: синтаза; под: +Fе2+) гем® Нb(над: +белок) Последняя реакция соединения гема с белком идет в цитозоле.]
После образования АЛК в митохондриях, в цитозоле происходит конденсация двух молекул АЛК с образованием порфобилиногена (монопиррола). Следующим этапом синтеза является реакция конденсации 4-х молекул порфобилиногена с образованием уропорфириногена (тетрапиррола).
В природе встречаются порфирины типов 1 и 3. В организме человека более широко представлены и биологически более важные изомеры 3 типа. В процессе декарбоксилирования уропорфобилиноген превращается в копропорфобилиноген, который из цитозоля поступает в митохондрии, где превращается в протопорфирин. Реакция окисления идет в присутствии кислорода при участии оксидаз. Заключительной стадией синтеза гема является включение 2-х валентного железа в протопорфирин. Эта реакция синтезируется гем–синтазой и завершает в митохондриях образование гема.
При нарушении синтеза гема в организме человека главным образом в печени могут развиваться порфирии – группа заболевании возникшие в результате блокирования отдельных стадий синтеза гема и сопровождающиеся увеличенным содержанием порфиринов в организме.
Причинами порфирий могут быть:
1. наследственные болезни, когда из-за дефицита митохондрий и ЦПМ ферментов синтеза гема, нарушается пигментный обмен;
2. приобретенные (токсические), которые вызываются действием ряда токсических соединений. Например: гексохлорбензол, соли тяжелых металлов, алкоголь. Эти соединения могут быть ингибированны ферментами в системе синтеза гема.
Порфирии могут сопровождаться поражением кожи под влиянием УФЛ, сопровождаясь фотодерматозами. Моча таких больных – розовая или красная.
Фотодерматозы – проявления кожной порфирии, т.е. увеличения светочувствительности кожи.
Распад гемаБольшая часть гемхромагенных пигментов в организме человека образуется при распаде гема. Главным источником гема является гемоглобин. В эритроцитах содержание гемоглобина составляет 80%, время жизни эритроцита 110-120 дней. В течение суток в организме взрослого человека массой 70 кг распадается около 6г гемоглобина. Распад эритроцитов происходит в клетках мононуклеарных фагоцитов. Наибольшее число таких клеток в селезенке, печени, костном мозге.
Первая реакция распада гемоглобина – это гидролиз его белковой части.
Катаболизм гема, освобождение от белковой части происходит в микросомальной фракции при участии сложной гем-оксигеназной ферментативной системы, требующей наличие НАДФН2 и кислорода. При поступлении гема в гемаксиназную систему микросом ионы железа окисляются в ферри форму, т.е. гем превращается в гемин.
гем (Fe2+)® (над: оксигеназа) гемин(Fe3+) (это ферри форма) ® (над: редуктаза; под: НАДФH2®НАДФ) гемин (Fe2+) (ферро форма) ® (над: редуктаза, +О2; под: НАДФН2®НАДФ)оксигемин (Fe3+)® (над: редуктаза, +О2; под: -СО, -Fe3+)биливердин® ( над: НАДФН2®НАДФ) ®билирубин® (над: +альбумин) билирубин-альбумин (неконъюгированный билирубин - НБ).
Как видно из схемы, в процессе реакций катаболизма гема, гемин восстанавливается с помощью редуктаз (НАДФН2) в ферро-форму (Fe2+).
При участии НАДФН2, кислород проходит последующее превращение с раскрытием тетрапирольного кольца гема, с выделением окиси углерода (СО) и освобождением ферри-иона (Fe3+).
Образуется ациклическое соединение биливердин, у которого редуктаза восстанавливает метиловый мостик между 3 и 4 пирролами, образуется желтый, токсический для клеток, пигмент – билирубин.
За сутки у взрослого человека массой 70 кг образуется 250-350 мг билирубина. Химические превращения гема в билирубин в клетках МФ можно наблюдать внутри организма в гематоме, где пурпурный цвет, обусловленный гемом, медленно переходит в желтый цвет, обусловленный билирубином.
Образованный в клетках билирубин является токсическим веществом, удаляется из них и поступает в кровь, взаимодействуя с транспортными белками - альбуминами. Образованное комплексное соединение билирубин-альбумин называется неконъюгированный билирубин - НБ.
НБ имеет свойства:
1. токсичен;
2. гидрофобен;
3. адсорбирован на альбумине;
4. не проходит через почечный эпителий;
5. не дает прямой реакции с диазо-реактивом Эрлиха.
НБ с помощью альбумина поступает для детоксикации в печень, где в гепатоцитах, в реакции конъюгации с глюкуроновой кислотой, при участии ТФ образуются:
- билирубин-моноглюкурониды (20%);
- билирубин-диглюкурониды (80%).
Эти билирубины носят название конъюгированного билирубина – КБ.
КБ имеет свойства:
1. не токсичен;
2. гидрофилен;
3. не связан с белками;
4. легко проникает через почечный барьер;
5. дает прямую реакцию с диазо-реактивом Эрлиха.
Этот билирубин (КБ) может проникать в кровяные капилляры. В плазме крови на его долю приходиться 25%, от общего билирубина, который в норме составляет 8-20 мкмоль/л.
Далее из печени КБ в составе желчи поступает в клетки, где под влиянием ферментов микрофлоры он гидролизуется. Отщепившаяся глюкуроновая кислота всасывается в слизистую кишечника и через воротную вену вновь поступает в печень, где может использоваться для детоксикации.
Билирубин под влиянием ферментов кишечной микрофлоры многократно восстанавливается, превращается в мезобилиноген, часть которого может всасываться слизистой и через систему воротной вены поступать в печень, где разрушается до моно-, дипирролов, которые из организма удаляются в составе желчи с каловыми массами.
Незначительная часть мезобилиногена с током крови поступает в почки, где превращается в другой пигмент – уробилиноген, который, окисляясь, образует пигмент мочи – уробилин.
В моче здорового взрослого человека массой 70 кг уробилин присутствует в следовых количествах.
За сутки (суточный диурез в норме равен 1,2-1,5 л) выделяет 1-4 мг уробилина.
Большая часть мезобилиногена в толстом отделе кишечника под влиянием ферментов кишечной микрофлоры, восстанавливаясь, превращается в стеркобилиноген – основной пигмент кала, который, окисляясь, превращается в стеркобилин. За сутки с калом выводиться примерно 280-300 мг стеркобилина.
Патология пигментного обменаКак правило, связана с нарушением процессов катаболизма гема и выражается гипербилирубинемией и проявляется в желтушечности кожи и видимых слизистых оболочек. Накапливаясь в ЦНС, билирубин вызывает интоксикацию. При гипербилирубинхмиях меняется соотношение НБ и КБ.
Выделяют следующие желтухи:
1. Механическая (обтурационная), которая связана с нарушением оттока желчи в клетках по причинам болезни печени, желчного пузыря, при закупорки желчевыводящих путей в случае глистной инвазии, при сдавлении опухолью из вне. В процессе застоя желчи растягиваются желчные капилляры, увеличивается их проницаемость.
КБ поступает в кровь, развивается гипербилирубиннемия, за счет КБ.
В крови резко увеличивается активность ферментов: щелочной фосфатазы, гамма-глутамилтрансминазы (ГГТ).
В моче резко увеличивается содержание билирубина, уменьшается уробилина. Количество стеркобилина в кале резко уменьшается, кал становиться ахоличным, т.е. обесцвечивается.
2. Паренхиматозная (острые гепатиты). Повреждение гепатоцитов при острых гепатитах (инфекционные заболевания, вирус гепатита, отравление), при этом уменьшается активность глюкоуронил-ТФ, что ведет к нарушению образования КБ. Токсическое воздействие на биомембрану ведет к увеличению проницаемости мембран гепатоцитов и переводу в кровь НБ и КБ. Развивается смешенная гипербилирубинемия.
В крови резко увеличивается активность ферментов: ЛДГ, АлАТ, АсАТ.
В моче, в больших количествах обнаруживается КБ – гипербилирунурия, уробилин, содержание стеркобилина в кале уменьшается.
3. Гемолитическая. Гемолиз эритроцитов может быть при гемолитических болезнях новорожденных, при переливании несовместимой группы крови, при дефиците ферментативных систем эритроцита, при гемоглобинозах (нарушение структуры гема, гемоглобина).
В крови наблюдается гипербилирубиннемия за счет НБ. Резко уменьшается количество эритроцитов и содержание гемоглобина.
В моче билирубин отсутствует, резко увеличивается содержание уробилина, содержание стеркобилина в кале увеличивается.
Моча таких больных интенсивно оранжевая, кал темный.
4. Физиологическая желтуха новорожденных. Обычно развивается на 3-4 сутки после рождения и держится в среднем 7-8 дней. Встречается у 80% новорожденных и связана с тем, что при рождении в крови содержится много эритроцитов, скорость распада которых высокая, а детоксикация билирубина в печени в первые дни после рождения развита слабо по причине низкой активности генов, ответственных за синтез глюкуронил-ТФ.
У недоношенных детей желтуха выражена сильнее и держится 4-5 недель. Если содержание билирубина в крови высокое и держится долго, то страдает, прежде всего, ЦНС. В этом случае необходимо проводить переливание крови, вводить барбитураты, которые индуцируют метаболизм билирубина в печени, т.е. конъюгацию с глюкуроновой кислотой. Проводимая светотерапия ускоряет выведение билирубина из организма.
5. Наследственная желтуха. Связана с врожденной недостаточностью (дефицитом) глюкуронил-ТФ. В крови у таких больных очень большое содержание билирубина за счет НБ. В желчи КБ нет. В отличие от гемолитической желтухи, количество эритроцитов и содержание гемоглобина в норме.
Биохимия крови Типы изменения биохимического состава кровиI. Абсолютные и относительные.
Абсолютные обусловлены нарушением синтеза, распада, выведения того или иного соединения.
Относительные обусловлены изменением объема циркулирующей крови (ОЦК). Когда ОЦК увеличивается (за счет воды), концентрация растворенных в ней соединений снижается. Если же ОЦК увеличивается (обезвоживание), то наблюдается сгущение крови и повышение концентрации веществ.
II. Качественные и количественные.
Качественные изменения: появление патологических компонентов (патологические белки – парапротеины) или отсутствие нормальных компонентов (афибриногенемия – отсутствие фибриногена, анальфалипопротеинемия – отсутствие α-ЛП).
Количественные – повышение или снижение показателя. Название образуется из приставки гипер- или гипо- соответственно, названия этого компонента и концовки -емия.
Белковый состав крови Функции белков крови:1. поддерживают онкотическое давление (в основном за счет альбуминов);
2. определяют вязкость плазмы крови (в основном за счет альбуминов);
3. определяют устойчивость форменных элементов в кровотоке (предупреждают склеивание клеток крови между собой);
4. участвуют в поддержании КЩС (кислотно-щелочного равновесия), образуя белковую буферную систему;
5. транспортируют метаболиты, биорегуляторы, микроэлементы, ксенобиотики (в основном за счет альбуминов). Напр., тироксинсвязывающий белок транспортирует гормон тироксин (Т4);
6. участвуют в регуляции гемостаза, являясь компонентами свертывающей и противосвертывающей систем;
7. участвуют в реакциях иммунитета (γ-глобулины, комплемент);
8. являются резервом аминокислот.
Общий белокВ норме общий белок крови 65-85 г/л.
Общий белок – это сумма всех белковых веществ крови.
► Гипопротеинемия – снижение альбуминов. Причины:
1. дефицит аминокислот в организме (из-за нарушения поступления, низкобелковой диете, нарушении переваривания и всасывания);
2. усиление распада белков (голодание, повышение потребности в энергии и строительном материале при беременности и травме);
3. выведение белков из кровеносного русла: выход в ткани (экссудация, транссудация) либо из организма – выход в мочу при нарушении фильтрации почками;
4. нарушение белоксинтезирующей функции печени.
► Гиперпротеинемия может быть в двух вариантах:
а) парапротеинемия – появление патологических белков;
б) повышение за счет белковой фазы воспаления.
АльбуминыВ норме 35-50 г/л.
Гипоальбуминемия – причины см. выше (гипопротеинемия). При снижении альбуминов ниже 30 г/л возникают отеки.
Гиперальбуминемия – при переливании альбуминов с лечебной целью и при сгущении крови (относительная гиперальбуминемия). Истинной не встречается.
Глобулины в норме 20-30 г/лI. α1 -глобулины
α-антитрипсин – ингибирует трипсин, пепсин, эластазу, некоторые другие протеазы крови. Выполняет антивоспалительную функцию крови. Повышенный уровень антитрипсина характерен для острой фазы воспаления, поэтому он называется белком острой фазы.
α1-серомукоиды – гликопротеины сыворотки крови. Являются компонентами клеточных мембран и появляются в крови вследствие обновления кклеток. Обнаруживаются в печени в ответ на воспаление (в любом органе). Повышается содержание серомукоидов в ответ на воспаление и процессы распада (при деструктивных процессах).
α1-фетопротеин – фетальный белок. В норме в больших количествах во внутриутробном периоде. Концентрация у взрослого человека не более 15мг/мл. Уровень фетопротеинов используется как маркерна уровне внутриутробного развития. Также используется как онкомаркер – маркер рака печени. В организме плода выполняет важные функции: заменяет альбумин, защищает от избытка материнского эстрогена, участвует в развитии печени.
II. α2 -глобулины
α-макроглобулин – защищает функционирующие белки от протеолитических ферментов. Это белки острой фазы воспаления. Их концентрация увеличивается при потере альбуминовой фракции с мочей, а также в острую фазу воспаления.
Гаптоглобин – связывает и транспортирует свободный гемоглобин в клетки ретикулярной системы. Сберегает железо (Fe), обезвреживает гемоглобин. Снижается гаптоглобин при гемолизе, повышается – в острую фазу воспаления.
Церрулоплазмин выполняет ряд функций:
1) транспортирует ионы меди (II) – связывает и удерживает Cu++ в кровеносном русле;
2) катализирует окисление Fe2+ в Fe3+;
3) обладает противовоспалительным действием;
4) является антиоксидантом – обезвреживает активные формы кислорода и ПОЛ (перекисное окисление липидов).
Повышение церрулоплазмина – при остром воспалительном процессе, циррозе печени, заболеваниях печени воспалительного характера.
Понижение – при нарушении его синтеза в печени (а также наследственные факторы).
Болезнь Вильсона-Коновалова: церрулоплазмин не удерживается в крови, а выходит в ткани и с мочой, а также откладывается в прозрачных средах глаза.
III. β-глобулины
Трансферрин – белок, транспортирующий ионы Fe3+. Повышение трансферрина – при дефиците железа в организме. Понижение – при остром воспалительном процессе (!) или при нарушении функций печени. У пациентов с врожденной гипо- или атрансферринемией не удается вылечить анемию.
Гемопексин – связывает и транспортирует гем в клетки ретикулоэндотелиальной системы. Повышение гемопексина – при гемолизе. Понижение – при остром воспалении.
Фибриноген (норма 2-4 г/л) – компонент свертывающей системы крови. При формировании сгустка превращается в фибрин. Фибриноген содержится в плазме крови, а в сыворотке крови его нет. Гиперфибриногенемия – при острой фазе воспаления и тромботических заболеваниях. Гипофибриногенемия – при нарушении синтеза его в печени (развивается гипокоагуляция).
С-реактивный белок – реагирует с С-полисахаридом пневмококка, реагирует с полисахаридами многих бактерий, клеточных стенок чужеродных клеток. Эта реакция необходима для фагоцитоза, следовательно, С-реактивный белок обладает противовоспалительным действием, способствует фагоцитозу, обеспечивает взаимодействие клеток иммунной системы. В норме содержится в количестве 5 мг/л. Повышение концентрации – при бактериальной инфекции и неинфекционных воспалениях – растет в тысячи раз, поэтому его называют маркером острого воспаления.
IV. γ-глобулины (иммуноглобулины, антитела)
Повышение титра иммуноглобулинов:
Ig M – острый инфекционный процесс;
Ig G – хронический воспалительный процесс и после острого заболевания;
Ig A – секреторный иммуноглобулин поверхности слизистых оболочек;
Ig E – антитела аллергии;
Ig D – его функция неизвестна.
Гипергаммаглобулинемия характеризует острый бактериальный воспалительный процесс или иммунопатологию. Обеспечивается плазматическими клетками.
Гипогаммаглобулинемия характеризует подавление звена гуморального иммунитета (при лучевой болезни, иммунодефиците, истощении организма).
Качественные изменения заключаются в появлении парапротеинов (продуктов аномального синтеза иммуноглобулинов), патологии клеток иммунной системы (их опухоли).
Напр., криоглобулин (чувствительный к холоду) переходит в гель при температуре ниже 37ºС (это обратимый процесс). Он обуславливает холодовую аллергию.
Небелковые азотсодержащие вещества Остаточный азотОстаточный азот – это сумма азота всех небелковых азотсодержащих веществ крови. В норме 14-28 ммоль/л.
1. Метаболиты:
1.1. аминокислоты (25%);
1.2. креатин (5%);
1.3. полипептиды, нуклеотиды (до 3,5%).
2. Конечные азотистые продукты:
2.1. мочевина (50%);
2.2. мочевая кислота (4%);
2.3. креатинин (2,5%);
2.4. индикан, аммиак.
Гиперазотемия (азотемия). Причины:
1) Продукционный фактор – вследствие распада белков и увеличенного содержания АК в составе остаточного азота.
Повышение аминокислот – гипераминоацидемия – при голодании, истощающих заболеваниях, гиперфункции щитовидной железы.
2) Ретенционный фактор – задержка азотистых шлаков в организме за счет нарушения функции почек. Напр., повышение мочевины, повышение креатинина (креатинин только фильтруется, но не реабсорбируется).
При интенсивном распаде нуклеиновых кислот, подагре повышается мочевая кислота.
При патологии мышц увеличивается креатин.
Углеводный обмен
Глюкоза в капиллярной крови натощак 3,3-5,5 ммоль/л.
1. Гипергликемия (повышение глюкозы):
1.1. панкреатическая гипергликемия – при отсутствии инсулина, нарушении функции В-клеток поджелудочной железы (сахарный диабет I типа – инсулинзависимый);
1.2. внепанкреатическая гипергликемия – при гиперпродукции контринсулярных гормонов, нарушении инсулиновых рецепторов (сахарный диабет II типа – инсулиннезависимый).
2. Гипогликемия (снижение глюкозы):
2.1. панкреатическая гипогликемия – инсулина (опухоль поджелудочной железы);
2.2. внепанкреатическая гипогликемия – развивается при недостатке контринсулярных гормонов (нарушении функции надпочечников, напр., их туберкулезе).
Кетоновые тела в норме не более 0,1 г/л. Представлены ацетоном, ацетоуксусной кислотой, бета-гидроксимасляной кислотой. Кетогенез усиливается при голодании, сахарном диабете, может приводить к кетоацидозу.
Липидный обмен
Холестерин в норме 3-5,2 ммоль/л. В плазме находится в составе ЛПНП, ЛПОНП (атерогенные фракции) и ЛПВП (антиатерогенная фракция). Вероятность развития атеросклероза связана с коэффициентом атерогенности:
К=(ЛПНП +ЛПОНП) /ЛПВП.
В норме у взрослого коэффициент атерогенности не более 3,0. У новорожденного – не более 1,0.
Гиперхолестеринемия может свидетельствовать об:
ü атеросклерозе (избыток ЛПНП и ЛПОНП);
ü сахарном диабете (избыток ацетил-Ко А);
ü ожирении;
ü гипотиреозе;
ü обтурационной (подпеченочной) желтухе.
Гипохолестеринемия:
ü голодание;
ü нарушение функции печени;
ü гипертиреоз (за счет катаболизма).
Билирубин общий в норме 8-20 мкмоль/л, конъюгированый – 2-8 мкмоль/л. Чаще встречается гипербилирубинемия.
Минеральный обменНатрий – это основной внеклеточный ион. На уровень Na+ в крови влияют минералокортикоиды (альдостерон задерживает натрий в почках). Уровень натрия увеличивается за счет гемоконцентрации, т.е. сгущения крови. Также увеличивается при избытке NaCl в пище.
Снижается уровень натрия при длительной рвоте (потеря бикарбонатов), длительных поносах (например, при холере) и при увеличении содержания воды в крови.
Калий – это главный внутриклеточный ион. Увеличивается его концентрация при задержке натрия в организме (например, при гиперсекреции альдостерона), при избытке в пище калийсодержащих продуктов (например, бананов), при нарушениях функций почек.
Снижается уровень калия при его недостатке в продуктах питания и при синдроме гемоделлюции. Снижение уровня калия ведет к нарушению сердечного ритма.
Кальций в норме 2,2-2,6 ммоль/л.
Гиперкальцийемия:
ü массивный распад костной ткани;
ü повышение секреции ПТГ (парат-гормон);
ü гиповитаминоз D;
ü снижение выработки кальциотонина.
Гипокальцийемия:
ü недостаточность паращитовидных желез;
ü гиперсекреция кальцитонина;
ü гиповитаминоз D.
Фосфор в норме 1-2 ммоль/л.
Гиперфосфоемия:
ü гипопаратиреоз;
ü гиповитаминоз D;
ü распад костной ткани.
Гипофосфоемия:
ü гиперпаратиреоз;
ü рахит;
ü нарушение всасывания фосфатов в ЖКТ.
Показатели КЩС крови в норме рН = 7,36 – 7,44.
Нарушения:
ü ацидоз – закисление крови (снижение рН);
ü алкалоз – защелачивание крови (повышение рН).
ВВ (buffer basis) – сумма гидроксилов всех буферных систем крови. В норме ВВ = 40 – 60 ммоль/л. Сдвиги КЩС, вызванные изменением показателя ВВ в отсутствии изменения рН, являются компенсироваными. Если же сдвиги сопровождаются изменением рН – декомпенсированые.
Ферменты плазмы кровиКлассифицируются:
1. Функционирующие ферменты (собственно плазменные). Напр., ренин (повышает АД через ангиотензин II), холинестераза (расщепляет ацетилхолин). Их активность выше в плазме крови, чем в тканях. Повышение или понижение их активности имеет значение в диагностике.
2. Нефункционирующие ферменты (плазмонеспецифические). Появляются в плазме крови из клеток тканей и органов. В крови не выполняют какой-либо функции, а выполняют ее в тканях. Это внутриклеточные ферменты. Их активность в плазме значительно ниже, чем в тканях. Напр., АлАТ, АсАТ, ГГТП (гамма-глютамил-транспептидаза), щелочная фосфатаза (ЩФ). Чаще наблюдается повышение их содержания, чем понижение.
Причины гиперферментемии:
1) повышение проницаемости биомембран;
2) цитолиз;
3) некроз тканей;
4) усиление синтеза.
Напр., ЩФ (щелочная фосфатаза) повышается при рахите (синтезируется остеобластами).
АлАТ (аланиновая аминотрансфераза) в норме 0,1-0,68 ммоль/ч·л. Наибольшее ее количество – в печени, меньше – в скелетных мышцах, др.
Повышение активности АлАТ при заболеваниях печени, скелетных мышц, миокарда. Это цитоплазменный фермент цитолиза. Активность его повышается в ранние стадии заболевания.
АсАТ (аспарагиновая аминотрансфераза) в норме 0,1-0,45 ммоль/ч·л. Наибольшее содержание в мышцах (скелетной, сердечной), в печени и др. АсАТ содержится не только в циотоплазме, но и в митохондриях, поэтому ее появление свидетельствует о более глубоком повреждении.
Повышение активности АсАТ – при заболеваниях мышц, генетических заболеваниях, травмах, воспалениях, а также при заболеваниях миокарда, печени. При заболевании печени повышение АсАТ, равное повышению АлАТ, означает глубокое повреждение.
ЛДГ (лактатдегидрогеназа) в норме не более 4 ммоль/ч·л. ЛДГ представлена пятью изоферментами:
ЛДГ1 – Н4 – только в миокарде;
ЛДГ2 – Н3М1 – в миокарде, эритроцитах, др.;
ЛДГ3 – Н2М2 – в легких;
ЛДГ4 – Н1М3 – в скелетных мышцах, печени;
ЛДГ5 – М4 – в печени.
КФК (креатинфосфокиназа) в норме 0,15-0,3 ммоль/ч·л. Содержится в мышечной ткани. Состоит из двух субъединиц: В (мозговой тип) и М (мышечный тип). Представлен тремя изоферментами:
КФК1 – ВВ – около 0% - в мозговой ткани;
КФК2 – МВ – менее 3% - в миокарде;
КФК3 – ММ – 97-100% - в скелетных и сердечной мышцах.
Активность КФК1 увеличивается при патологии нервной системы с явлениями деструкции нервной ткани (например, опухоль, травма, ушиб мозга). Активность КФК2 при поражениях миокарда (например, инфаркт). КФК3 – при поражениях миокарда и скелетных мышц (например, при синдроме длительного сдавливания).
ЩФ (щелочная фосфатаза) в норме 0,5-1,3 ммоль/ч·л. Источники ЩФ:
1) костная ткань (синтезируется остеобластами);
2) желчевыводящая система (синтезируется эпителием желчных протоков).
ЩФ повышается при заболеваниях костной системы, метастазах опухолей в костную ткань, рахите, остеопорозе, подпеченочных (обтурационных) желтухах.
α-амилаза синтезируется в поджелудочной железе и в слюнных железах. В крови повышается при панкреатитах, реже – при воспалении слюнных желез. Панкреатическая амилаза фильтруется в мочу.
Биохимия почек
Функции почек
I. Экскреторная функция – выделение мочевины, мочевой кислоты, креатинина, ксенобиотиков, лекарств, токсинов, избытка воды, микроэлементов, электролитов.
1. Ультрафильтрация – в клубочках за счет разницы гидростатических давлений образуется первичная моча (180 л/сут), которая по своему составу почти не отличается от плазмы крови, но практически не содержит белка (могут присутствовать микроколичества альбуминов). Т.е. образуется безбелковый фильтрат.
2. Канальцевая реабсорбция – обратно всасывается вода, ионы натрия, хлора, магния, фосфора, гидрокарбонаты, глюкоза, белок, т.е. те вещества, в которых нуждается организм. В результате моча становится более концентрированной. Способность концентрировать мочу – важный показатель функции почек. Он оценивается по плотности мочи.
Практически не реабсорбируется креатинин, в небольшом количестве реабсорбируется мочевина, мочевая кислота, т.е. имеет место избирательная реабсорбция.
3. Секреция ионов калия, аммония, водорода. При увеличении концентрации веществ секретируется вода. Образуется вторичная моча.
Вещества, находящиеся в моче, делятся на:
1) в норме не фильтруемые грубодисперсные высокомолекулярные соединения, которые в норме не могут преодолеть почечный барьер (глобулины). Их появление говорит о нарушении фильтрации.
2) в норме фильтруемые: присутствуют в первичной моче, но не все из них присутствуют во вторичной моче. Они делятся на пороговые и беспороговые.
Пороговые из первичной мочи реабсорбируются. Это аминокислоты, глюкоза. Во вторичной моче в норме не содержатся. Имеется порог реабсорбции.
Беспороговые выделяются во вторичную мочу независимо от функции почек: мочевина, креатинин. Они не подвергаются реабсорбции.
Порог реабсорбции – верхний предел концентрации вещества в крови, преодоление которого влечет неэффективное обратное всасывание. Для глюкозы -–10 ммоль/л.
Появление пороговых веществ в крови влечет появление их в моче.
II. Регуляторно-гомеостатическая: регуляция АД (альдостерон, ренин), объема внеклеточной жидкости (регуляция гормонами, для которых орган-мишень – почки: вазопрессин, альдостерон), регуляция осмотического давления (вазопрессин, альдостерон). Также регулируется кислотно-щелочное равновесие.
Почки способны секретировать избыток протонов (Н+) в мочу – ацидогенез. Он происходит так:
1) под влиянием карбоангидразы образуется угольная кислота (СО2 + Н2О → Н2СО3);
2) угольная кислота распадается на протон (выделяется в мочу) и гидрокарбонат-анион (идет в кровь) (Н2СО3 → НСО3-- (в кровь) + Н+ (в мочу));
3) в моче протоны взаимодействуют с гидрофосфатом натрия с образованием дигидрофосфатов. (Н++Na2HPO4® Na++NaH2PO4) Освободившийся ион натрия реабсорбируется в кровь, где соединяется с гидрокарбонат-анионом (НСО3-- +Na+®NaНСО3). Т.о., натрий сберегается, а протоны выводятся.
Ацидогенез обеспечивает:
а) выведение избытка протонов,
б) сбережение ионов натрия и гидрокарбонатов, благодаря чему поддерживается буферная ёмкость крови.
Кроме того, почками в мочу выделяются летучие жирные кислоты, аммиак – аммониогенез.
Аммониогенез – более простой процесс, чем ацидогенез. Происходит в эпителии почечных канальцев:
1) Аммиак доставляется в клетку в составе глутамина (ГЛН). ГЛН под действием глутаминазы распадается на глутаминовую кислоту (ГЛУ) и аммиак. Кроме того, аммиак может самостоятельно поступать в клетку.
2) Аммиак выводится в мочу, где взаимодействует с протоном. В результате образуется катион аммония (H++NH3® NH4+). Этот катион взаимодействует с анионом хлора с образованием нейтральной соли (NH4++Cl--® NH4Cl).
В итоге:
а) выводится аммиак;
б) связываются протоны в моче и уменьшается ее кислотность;
в) выводятся хлориды.
III. Антитоксическая функция. Реакция конъюгации токсических веществ с глицином, уксусной и глюкуроновой кислотами. Реакции окисления некоторых токсических веществ.
IV. Внутрисекреторная функция (ренин, эритропоэтин – стимулирует красный росток кроветворения, простагландины). Часть клеток почек рассматриваются как эндокринные.
Физические свойства мочи здорового человека, их изменения при патологии
I. Количество мочи в норме 1,2-1,5 л.
► Полиурия – увеличение количества мочи из-за:
1) увеличения фильтрации (под действием адреналина увеличивается фильтрационное давление в капиллярах почек);
2) увеличения онкотического давления при снижении концентрации белков в плазме;
3) снижения реабсорбции воды и натрия при недостатке вазопрессина (несахарный диабет);
4) снижения активности натрий-сберегающих процессов (ацидогенез и аммониогенез);
5) повышения концентрации осмотически активных веществ (глюкоза, белки, мочевина и др.);
6) угнетения систем реабсорбции при повреждении почечных канальцев.
► Олигоурия – снижение количества мочи из-за:
1) снижения фильтрации (снижение АД и давление в капиллярах почечных клубочков);
2) снижения онкотического и коллоидно-осмотического давления крови (парапротеины);
3) снижения обратного всасывания натрия и воды (гиперсекреция альдостерона и вазопрессина);
4) угнетения всех фаз образования мочи (резкое снижение количества функционирующих нефронов).
► Анурия – полное отсутствие мочи при выраженной почечной недостаточности.
II. Цвет в норме от светло-желтого до темно-желтого.
При патологии моча может быть:
ü бесцветна за счет полиурии;
ü насыщенно-желтого цвета при обезвоживании;
ü цвета пива (коричневато-зеленовато-бурый) за счет желчных пигментов при подпочечной и печеночной желтухах из-за появления конъюгированого билирубина;
ü розового цвета при эритроцитурии (в умеренном количестве) или при патологии почечного фильтра;
ü ярко-красного цвета при наличии в моче свежей крови (травма мочевыводящей системы или гематурия, которая может быть почечной и внепочечной).
III. Запах в норме специфический. При патологиях может быть:
ü резко-аммиачный при повышенном распаде белков в организме;
ü фруктовый или ацетона – при наличии кетоновых тел (сахарный диабет, голодание);
ü гнилостный – процессы распада в мочевом пузыре (гангрена);
ü каловый – при наличии пузырно-ректальных свищей.
IV. Реакция среды в норме слабо-кислая (рН=5,0-7,0).
ü Увеличение рН (щелочная реакция) – угнетение функции почек, алкалоз.
ü Понижение рН (кислая реакция) – ацидоз за счет удаления избытка протонов и за счет выделения кетоновых тел.
При употреблении значительного количества белковой пищи происходит закисление, а растительной – защелачивание.
V. Суточные колебания плотности в норме 1,005-1,030 г/мл.
Одинаково монотонная плотность мочи – изоастенурия. При нарушении концентрирующей способности почек может быть:
ü понижение плотности – гипостенурия, преобладание процессов фильтрации над реабсорбцией (сопровождается полиурией). Возникает при несахарном диабете и заболеваниях почек (уменьшается синтез вазопрессина);
ü повышение плотности – гиперстенурия при появлении веществ, в норме не содержащихся: глюкоза, кровь, белки, избыток слизи в моче.
VI. Прозрачность мочи в норме полная. Может быть:
ü мутная из-за присутствие нерастворимых солей (уратов, оксалатов), слизи, гноя, белка.
Показатели химического состава мочи
Общий азот – это совокупность азота всех азотсодержащих веществ в моче. В норме – 10-16 г/сутки. При патологиях общий азот может:
ü увеличиваться – гиперазотурия – из-за распада белков в организме или употребления большого количества мясной пищи;
ü уменьшатся – гипоазотурия – при нарушении функции почек.
В состав общего азота входят:
I. Мочевина в норме 20-35 г/сут. Может быть:
ü повышение мочевины – при повышенном распаде белков в случае нормальной функции печени и высокобелковом питании;
ü понижение мочевины – при нарушении синтеза в печени и нарушении функции почек.
II. Мочевая кислота в норме 0,3-1,2 г/сут. Она является продуктом катаболизма пуринов. Может быть:
ü гиперурикурия – при распаде нуклеопротеинов, употреблении большого количества белковой пищи, некоторых видах подагры;
ü гипоурикурия – при определенной форме подагры с нарушением выведения мочевой кислоты, нарушении ее фильтрации почками.
III. Креатинин в норме 0,8-2,3 г/сут. Может быть:
ü повышение креатинина – при усиленном распаде компонентов мышечной ткани, в случае тяжелой мышечной работы, травмы мышц, при увеличении потребления мясной пищи;
ü понижение – при угнетении фильтрации почками.
IV. Белок в норме не больше 30 мг/л. Качественными реакциями не определяется. Может быть:
ü протеинурия при гематурии или при нарушении функции почек.
Патологические компоненты
В норме они в моче отсутствуют.
Желчные пигменты, билирубин - при их появлении можно диагносцировать почечную, механическую желтухи.
Кетоновые тела – при их появлении сахарный диабет или голодание.
При появлении глюкозы наблюдается глюкозурия, что говорит о сахарном диабете и почечной недостаточности.
При появлении крови можно говорить о повреждении мочевыводящих путей или почечной недостаточности.
Индолурия в отсутствии индикана говорит об угнетении обезвреживающей функции печени.
Индиканурия в отсутствии индола говорит о повышенных процессах гниения белков в кишечнике в отсутствии нарушения функции печени.
Биохимия нервной ткани
Химические компоненты нервной ткани
ЦНС координирует и регулирует обмен веществ в организме. Она также обеспечивает взаимосвязь организма с внешней средой.
Химический состав нервной ткани сложен и неоднороден. Например, в сером веществе 77-81% воды, а в белом – 70%. Количество белков в нервной ткани меньше, чем в мышечной или в печени. При этом белков больше в сером веществе, и меньшее их количество содержится в периферической нервной ткани. В функционально более активных структурах белков больше.
Характерным для белков нервной ткани является то, что они находятся в комплексе с другими соединениями, т.е. это сложные белки. Больше всего липопротеинов (ЛП). Особенно много их в миелиновых оболочках. Есть фосфопротеины – фосфат присоединяется к белку через серин (-NH-CH(CH2OPO3H2)-CO-). Также в нервной ткани есть нуклеопротеины (НП) (дезоксирибонуклеопротеины (ДНП), рибонуклеопротеины (РНП)), гликопротеины (например, нейрокератин).
Особый интерес при изучении нервной ткани имеет исследование нейроспецифических белков:
1. Белок S-100 – растворим в 100% (NH4)2SO4. Обнаружен он в основном в нервной ткани. Принимает участие в процессах формирования памяти (при обучении животных его содержание возрастает). Есть предположение, что этот белок играет роль при возникновении наркотической зависимости.
2. Белок 14-3-2.
Пептиды, которые специфичны для нервной ткани, называются нейропептиды. Их около 100. 80-90% нейронных контактов осуществляются нейропептидами. К ним относятся пептиды памяти (аргинин-вазопрессин, лейцин-вазопрессин, окситоцин, АКТГ), пептиды боли (вещество Р, эндорфины, энкефалины и т.д.), пептиды сна.
Небелковые азотистые соединения. В нервной ткани количество азотистых соединений такое же, как и в других тканях, но некоторых больше:
1. свободных АК, особенно ароматических и дикарбоновых АК. Больше АК - предшественников нейромедиаторов (ТИР, ТРИ, АСП, ГЛУ);
2. циклических нуклеотидов (цАМФ, цГТФ).
Углеводы нервной ткани. Их запасы небольшие. Откладываются в виде гликогена, которого в нервной ткани 0,1% (в печени – 5-10%, в мышечной ткани – 0,2-2%). Глюкозы содержится 1-4 ммоль/кг.
Липиды. В нервной ткани их содержится достаточно много (в сером веществе – 25%, в белом – до 50%). Около 50% всех липидов представлены фосфолипидами (ацетальфосфатиды, плазмогены). Имеются гликолипиды: цереброзиды, ганглиозиды (их нет в других тканях). Жирные кислоты. В нервной ткани эти кислоты содержат большое количество двойных связей (4-5). 25% всех липидов составляет холестерол. В нервной ткани он находится в свободном виде. Нейтральные жиры содержатся в небольших количествах в ткани головного мозга, больше их в периферических нервах.
Минеральные вещества. Внутри клеток содержатся ионы калия, а в межклеточном пространстве – ионы натрия. Они участвуют в передаче нервного возбуждения. Также содержаться ионы кальция, магния, железа, меди, алюминия, цинка; анионы представлены фосфатами и АК белков.
Особенности обмена веществ в нервной тканиЭнергетический обмен.
В ткани головного мозга увеличено клеточное дыхание (преобладают аэробные процессы). Мозг потребляет большее количество кислорода, чем постоянно работающее сердце, в 20 раз больше, чем покоящиеся мышцы. 20-25% всего кислорода приходится на долю головного мозга. У детей до 50%.
Ткань головного мозга использует весь кислород, находящийся в ней, за 10 секунд. Следовательно, важное значение имеет кровоснабжение головного мозга. при нарушении кровообращения через 6-8 секунд наступает потеря сознания.
Дыхательный коэффициент (отношение объема СО2 к объему О2) в тканях головного мозга приблизительно равно 1, следовательно углеводы – это основной субстрат для окисления. Мозг – единственный орган, который использует в качестве источника энергии практически одну только глюкозу (при патологии могут использоваться кетоновые тела), т.е. функционирование головного мозга зависит от снабжения глюкозой.
70% АТФ в тканях головного мозга используется для поддержания ионных градиентов (энергия используется для удаления ионов натрия из клетки).
Углеводный обмен.
Исходным субстратом для окисления является глюкоза (не гликоген!). Гипогликемия приводит к судорогам и, возможно, к смерти.
85% глюкозы окисляется аэробно (до углекислого газа и воды), 15% - анаэробно (до лактата). Анаэробное окисление – это аварийный механизм.
Гликогена содержится немного – 0,1%, но интенсивность его обновления достаточно велика. Весь гликоген в ткани головного мозга обновляется за 4 часа. Распад гликогена идет 2 путями:
- фосфорилический (с участием фосфорилазы);
- гидролитический - g-амилаза отщепляет остатки глюкозы.
Нарушения обмена углеводов ведут к нарушению функций головного мозга. При авитаминозе В1 нарушается превращение ПВК, следовательно развиваются полиневриты. Угнетение окисления углеводов ведет к развитию торможения в нервной системе (используется при разработке снотворных веществ). Во сне потребление глюкозы снижается, а при возбуждении увеличивается.
Белковый обмен.
При возбуждении увеличивается распад белков и, как следствие, образуется больше аммиака и азота АК. При торможении распад белков снижается.
У человека в больших количествах образуется аммиак, являющийся токсичным веществом для нервной ткани и поэтому он должен быть обезврежен. Обезвреживание происходит путем образования амидов моноаминодикарбоновых АК: [рис. NH2-CH(CH2-CH2-COOH)-COOH (это глутаминовая кислота) +NH3® (над стрелкой глутамин-синтетаза, под Mg2+, АТФ®АДФ+Фн) NH2-CH(CH2-CH2-CONH2)-COOH (это глутамин)]. Этот процесс интенсивно протекает в нервной ткани, т.к. глутамин свободно выходит из клеток.
Глутаминовая кислота играет особенную роль в обмене веществ:
1. связывает аммиак;
2. участвует в реакциях переаминирования, в результате которых образуются заменимые АК (аспарагиновая кислота);
3. подвергается декарбоксилированию: [рис. NH2-CH(CH2-CH2-COOH)-COOH (это глутаминовая кислота) ® (над стрелкой глутамат-декарбоксилаза, под – ПФ(В6)) NH2-CH2-CH2-CH2--COOH (это g-аминомасляная кислота)]. Образующаяся g-аминомасляная кислота является тормозящим нейромедиатором;
4. подвергается окислительному дезаминированию. В результате этого многие АК теряют NH2-группу;
5. является возбуждающим нейромедиатором;
6. стабилизирует содержание ионов калия в клетках нервной ткани.
До 10% глюкозы используется в качестве субстрата для синтеза глутаминовой кислоты.
Липидный обмен.
В нервной ткани липиды не играют энергетической роли. Содержащиеся в основном фосфолипиды и холестерин играют структурную функцию. Нейтральные жиры играют защитную функцию.
Химическая передача нервного возбужденияПередача возбуждения с одной клетки на другую происходит с помощью нейромедиаторов:
- нейропептидов;
- АК;
- ацетилхолина;
- биогенных аминов (адреналин, норадреналин, ДОФА, серотонин).
Также в механизме передачи нервного возбуждения важную роль играют:
- натриевый насос (Na-K-АТФаза);
- натриевые каналы;
- калиевые каналы.
Последовательность процессов:
1. в результате воздействия раздражителя в синаптическую щель из визикул высвобождается нейромедиатор;
2. нейромедиатор диффундирует к мембранам 2 нервных клеток;
3. присоединяется к своему рецептору;
4. изменяется конформация рецептора;
5. происходит открытие натриевых и калиевых каналов, при этом ионы натрия идут в клетки, а калия - из них.
После удаления (разрушения) нейромедиатора начинает работать Na-K-насос, т.е. АТФаза удаляет ионы натрия из клеток, а ионы калия возвращаются. В результате очаг возбуждения снимается.
Каждый нейромедиатор действует в синапсах на свой рецептор. В холинергических синапсах основным медиатором является ацетилхолин (АХ). Он образуется из Ац-КоА и холина: [рис. (CH3)3-N-CH2-CH2OH (это холин) + CH3-COSKoA ®(холинацетилтрансфераза, -HSKoA) (CH3)3-N-CH2-CH2OСОСН3 (это АХ)]. Разрушается АХ под влиянием холинэстеразы.
В адренергических синапсах образуются ДОФамин, норадреналин. Образование происходит из фенилаланина, который сначала преобразуется в тирозин: ФЕН®(фенилаланингидроксилаза, +1/2О2) ТИР. Далее ТИР®(гидроксилаза, +1/2О2) ДОФА® (декарбоксилаза, -СО2) ДОФамин® (гидроксилаза, +1/2О2)норадреналин. [рис. всех этих формул] Разрушаются эти нейромедиаторы под действием моноаминооксидаз.
В серотонинергических синапсах образуется серотонин из АК триптофана: [рис. триптофан®(гидроксилаза, +1/2О2) 5-окситриптофан ® (декарбоксилаза, -СО2) 5-окситриптамин (серотонин)]. Разрушается под действием моноаминооксидаз
Биохимия мышечной ткани
Мышечная ткань составляет 40-42% от массы тела и около 50% от обмена веществ приходится именно на мышечную ткань, а при интенсивной мышечной работе: до 80% от обмена веществ.
Функции мышечной ткани:
1. сократительная;
2. теплопродукционная.
Структурные элементы: мембрана – сарколемма, цитоплазма – саркоплазма, сократительные элементы – миофибриллы. Строение под электронным микроскопом: изотропные диски, анизотропные диски, Z-линия.
Химический состав:
70-80% воды, 17-21% белков. Белки различны в разных местах: белки стромы – опорные (коллаген, эластин), белки саркоплазмы – ферменты (альбумины, глобулины, миоглобин). Миоглобин – хромопротеин, по структуре похож на гемоглобин и точно так же связывает кислород, но не трансформирует его. Белки миофибрилл: миозин (50% всех белков миофибрилл), актин (25% белков), тропомиозин, тропонины, актинины (все вместе составляют 25% от всех белков миофибрилл).
МиозинОтличается большим содержанием глутаминовой кислоты. Имеет отрицательный заряд. Он связывает ионы Са++ и Mg++. В присутствии Са++ миозин обладает активностью АТФ-азы. В присутствии Mg++ миозин связывает АТФ и АДФ. Способен взаимодействовать с актином.
Молекула миозина длинная – 160 нм и тонкая (ширина её – 2 нм), представляет собой две полипептидные цепи. Есть т.н. головки миозина.
АктинИмеет три формы.
- мономерная форма: G–актин (глобулярная структура, глобулы полярные), связывается с АТФ;
- димерная;
- полимерная.
Мономеры могут соединяться в присутствии АТФ в димеры (G+G+АТФ®G-АТФ-G+ Фн), из димеров могут образовываться полимеры: F–актин (от «фибрилла»).
ТропомиозинПредставляет собой две a- спирали (своеобразные нити). Он соединяется с тропонином и этот комплекс присоединяется к актину.
Тропонины
Глобулярный белок нескольких видов. Выделяют: TN-C, TN-I, TN-T.
TN-T связывается с тропомиозином, TN-I – ингибитор АТФ-азы, TN-C связывается с Ca++.
Имеют специфические отличия по АК составу от белков сердечной мышцы. Их можно определить с помощью иммуноферментного анализа.
Небелковые азотистые комплексы. Экстрактивные вещества
Экстрактивные вещества – небелковые азотистые вещества. К ним относятся:
АТФ: 0,25-0,40%.
Креатин-фосфат - 0,4-1,0% и его уровень растет при физической нагрузке. Он синтезируется из АРГ, МЕТ, ГЛИ, и может переходить в креатин. [рис. формулы креатин-фосфата COOH-CH2-N(CH3)-C(NH)-NH~PO3H2]
Карназин: b-аланилгистидин, участвует в транспорте фосфатных остатков. [рис. формулы карназина]
Ансерин отличается от карназина тем, что имеет метильную группировку; его функция - транспорт ионов Са++.
Карнитин: производная g-амино-b-гидроксимасляной кислоты. Корнитин транспортирует жирные кислоты через мембрану в митохондрии, поэтому мышечная ткань может использовать жирные кислоты в качестве источника энергии.
В мышечной ткани есть свободные АК, пуриновые основания, мочевина.
Углеводы мышечной ткани
Гликоген: 0,2-2%, но мышечная масса настолько велика, что содержание гликогена в мышцах в целом в 2 раза больше, чем в печени. Также содержаться гексозомонофосфаты, триозомонофосфаты, ПВК, молочная кислота, следы глюкозы (свободной почти нет).
Липиды – около 1%. Представлены нейтральными жирами в соединительно-тканных волокнах. Холестерол и фосфолипиды – компоненты биомембран. Жирные кислоты играют особую роль в миокарде как источник энергии.
Минеральные вещества. K+, Na+ участвуют в передаче возбуждения; также содержаться Ca++, Mg++, Fe++ (особенно много в миоглобине).
Химический состав мышечной ткани может изменяться при патологиях: мышечные дистрофии, полимиозиты, атрофия мышц. Всё это приводит к снижению фибриллярных белков и увеличению содержания белков стромы, саркоплазмы, снижению уровня АТФ, креатин-фосфата. Креатинурия развивается при многих мышечных патологиях, нарушается удержание его в мышечной ткани. В норме креатин образуется в печени из ГЛИ, МЕТ, АРГ, потом попадает в кровь, достигает мышцы, где превращается в креатин–фосфат при участии АТФ; часть креатина может превращаться в креатинин, который выводится с мочой. При патологии креатин из крови сразу поступает в мочу – креатинурия.
Энергетическое обеспечение мышечного сокращения
Есть некоторые особенности:
1. энергия необходима периодически;
2. при сокращении мышцы нарушается ее кровоснабжение.
Механизмы преодоления:
1. мышечная ткань содержит миоглобин, который связывает кислород;
2. мышечная ткань отличается большим содержанием фосфорорганических соединений, что позволяет без окислительного распада углеводов сокращаться мышце.
Запасы АТФ небольшие, они используются за 0,5 сек сокращения, но при этом уровень АТФ не снижается, т.к. есть механизмы ресинтеза АТФ.
Ресинтез АТФ – образование АТФ из АДФ и неорганического фосфата.
Пути ресинтеза АТФ:
1. креатинкиназная реакция:
[креатинфосфат+АДФ« (креатинфосфокиназа) креатин+АТФ] Это основная реакция, причем реакция будет идти вправо при сокращении, а влево - в состоянии покоя. Т.о., креатинфосфат играет роль транспорта энергии из митохонрий.
2. аденилаткиназная реакция:
АДФ+АДФ ®(аденилаткиназа) АТФ+АМФ. При этом АМФ подвергается распаду и выводится как мочевая кислота.
3. анаэробное окисление, при этом образуется молочная кислота. Эта реакция характерна для быстрых белых мышц.
4. аэробное окисление. Оно наиболее эффективно. Происходит окисление углеводов до воды и углекислого газа. Этот процесс характерен для красных мышц, окислению могут подвергаться не только углеводы, но и жирные кислоты.
Нарушения метаболизма при ишемической болезни сердца
Ишемия – недостаток кровоснабжения, при этом снижается уровень кислорода, снижается окислительное фосфсрилирование, увеличиваются анаэробные процессы. Это всё приводит к снижению уровня гликогена и повышению уровня лактата, что приводит к развитию ацидоза. Это в свою очередь приводит к ингибированию активности фосфофруктокиназы (блокируется гликолиз), снижается уровень АТФ, снижается уровень креатинфосфата. Нарушается проницаемость мембран, калий выходит из клетки, происходит выход ферментов в кровь (ферментемия). Содержание фибриллярных белков падает, увеличивается количество белков стромы. Нарушается окисление жиров в миокарде и вызывается жировая инфильтрация миокарда.
Биохимия мышечного сокращения
Раньше мышечное сокращение представляли как изменение структуры белка. Но эти представления были опровергнуты с помощью электронной микроскопии. Теория Хэнсона и Хаксли: укорочение за счет проникновения нитей актина между нитями миозина.
При этом необходимо наличие Са2+. В покое кальций находится в трубочках саркоплазматического ретикулума. Са-зависимая АТФ-аза как бы закачивает Са2+ в трубочки за счет распада АТФ. Т.е. если много АТФ, то свободного Са++ мало.
Если возникает раздражение нервного волокна, то Са2+ выходит из саркоплазматического ретикулума за счет изменения проницаемости мембраны, и выход его приводит к взаимодействию головки миозина с актином. Если есть Са2+ и АТФ, то белки продвигаются друг между другом. В покое миозин связан с Mg и АТФ. Если нет расщепления АТФ, то спайки не образуются. Выход Са2+ вызывает распад АТФ и образование спаек.
1. SR-Ca2+ ®(нервное возбуждение) SR+ Ca2+
2. активация актина: А-Тр+ Ca2+ ®А+ Тр- Ca2+
3. активация АТФ-азы кальция: Миозин-АТФ +Н2О+Са2+®(АТФ-аза) Миозин~Фосфат + АДФ
4. взаимодействие миозина и актина: М~Ф + А ®(+Са2+, +Н2О)М~А + Фн
5. М~А ®(сокращение)М-А+ работа
Для расслабления тоже нужна энергия АТФ:
1. М-А + АТФ®М-АТФ + А
2. связывание Са2+: Т-Са2++SR+АТФ®Т+SR- Са2++ АДФ+Фн
3. связывание тропонина с актином: Т+А®Т-А
Биохимия соединительной ткани
Соединительная ткань составляет 50% массы тела человека. Широко представлена в организме; это лимфоидная, жировая, костная ткани. Есть 3 принципа, по которым определяют соединительную ткань:
1. большое количество межклеточного вещества;
2. в межклеточном веществе присутствуют фибриллярные волокна (коллагеновые, эластиновые, ретикулярные);
3. главная функция заключается в синтезе комплексных веществ на экспорт (экстрацеллюлярные компоненты).
Функции соединительной ткани:
1. опорная (костная, хрящевая такни, сухожилия);
2. барьерная – связана с положением соединительной ткани. Защищает от проникновения инфекционных заболеваний Имеются процессы фагоцитоза и иммуногенеза;
 3. метаболическая –
синтезирует на экспорт белки, макромолекулярные вещества (коллагеновые
волокна), протеогликаны. Кортизол в фибробластах превращается в 11-b-оксиандростедион, который противоположен ему
(кортизолу) по действию. Так, кортизол угнетает пролиферацию и синтетическую
активность соединительной ткани, а 11-b-оксиандростедион
увеличивает анаболизм;
3. метаболическая –
синтезирует на экспорт белки, макромолекулярные вещества (коллагеновые
волокна), протеогликаны. Кортизол в фибробластах превращается в 11-b-оксиандростедион, который противоположен ему
(кортизолу) по действию. Так, кортизол угнетает пролиферацию и синтетическую
активность соединительной ткани, а 11-b-оксиандростедион
увеличивает анаболизм;
4. депонирующая – выполняет жировая ткань, в которой депонируются жиры;
5. репаративная функция – образование рубцовой ткани.
Клеточные элементы соединительной ткани:
1. фибробласты – продуцируют коллаген, эластин, гликозаминогликаны, протеогликаны;
2. тучные клетки (гепариноциты) – продуцируют гепарин, гистамин, 5-окситриптамин;
3. макрофаги;
4. плазматические клетки;
5. клетки, проникающие в соединительную ткань из крови (лимфоциты и др.).
Межклеточный матрикс соединительной ткани характеризуется наличием волокнистых структур.
Коллаген – наиболее распространенный белок (25-30% от всех белков человека). Более 80% всех белков он составляет в коже, костях, связках, сухожилиях, хрящах. Поэтому он долгое время считался белком соединительной ткани.
Коллаген характеризуется особым АК составом:
- 1/3 всех АК остатков приходится на глицин;
- значительное количество пролина (до 10%);
- встречается гидроксипролин и гидроксилизин.
[рис. 4-гидроксипролина и 5-гидроксилизина]
Большая часть представлена триадами –ГЛИ-Х-Y-, где Х – чаще пролин, а Y – чаще гидроксипролин. Эта регулярная последовательность представлена левозакрученной коллагеновой спиралью, более вытянутой, чем a-спираль. Каждая из спиралей представляет собой полипептидную цепь. Несколько спиралей соединяются в одну суперспираль, удерживающуюся за счет водородных связей между субъединицами. Длинна суперспирали примерно 300 нМ.
По АК составу выделяют 2 вида коллагеновых цепей:
- a1;
- a2.
a1 могут быть 4-х типов: a1(I), a1(II), a1(III), a1(IV).
Наиболее распространен 1 тип, куда входит и [a1(I)]2a2.
Процесс синтеза коллагена можно разделить на несколько этапов:
1. трансляция;
2. котрансляционная модификация цепи;
3. трансмембранный перенос;
4. внеклеточная модификация и образование коллагеновых волокон.
[рис. проколлагена: слева (N-конец) сигнальный пептид, затем 1050 АК остатков будущего коллагена, справа (С-конец) С-концевой пептид. Рисуется просто линия и на ней отмечается где и что].
Препроколлаген претерпевает процессинг в ходе прохождения через ЭПС и комплекс Гольджи до появления во внеклеточном пространстве. При этом происходят следующие процессы:
- гидроксилирование [рис. пролилпептид (в составе белка)+ a-КГ+ О2®(пролилгидроксилаза, витамин С) гидроксилпролилпептид + СООН-СН2-СН2-СООН (это сукцинат) +Н+. Пролилпептид рисуется как структура пролина с незакрытыми связями.]
- гликозимирование – внедрение углевода, возможно только после гидроксилирования [рис. гидроксилизин (с незакрытыми связями) +УДФ-галактоза® (галактозилтрансфераза) галактозилпропептид+ УДФ]. Гликозимирование препятствует действию протеаз и способствует возникновению межцепочечных водородных связей.
- формирование тройной спирали коллагена. После этого невозможно ни гидроксилирование, ни гликозимирование.
 |
|||
 |
 (zip - application/zip)
(zip - application/zip)