Министерство образования и науки РФ ФГБОУ ВПО
«Воронежский государственный лесотехнический университет им. Г.Ф.Морозова»
Кафедра ботаники и физиологии растений.
КУРСОВАЯ РАБОТА
ПО ДИСЦИПЛИНЕ « ФИЗИОЛОГИЯ РАСТЕНИЙ»
НА ТЕМУ « ВОДНЫЙ РЕЖИМ СОСНЫ КРЫМСКОЙ »
Воронеж 2016
Содержание
Введение…………………………………………………………………….3-4
1. Общая часть………………………………………………………………5-23
1.1. Обзор литературы……………………………………………………...5-15
1.2. Объект исследования и его характеристика…………………………16-20
1.3. Методика исследования……………………………………………….21-23
2. Специальная часть………………………………………………………24-29
2.1.Результаты исследований……………………………………………... 24-29
Выводы………………………………………………………………………30
Список использованной литературы……………………………………...31
Введение.
Физиология растений – это наука о процесса жизнедеятельности, происходящих в растении.
Как любая наука физиология растений имеет свой объект исследований, цель и задачи, методы исследований.
Объектом исследований физиологии растений является сам растительный организм.
Цель физиологии растений как науки – раскрыть сущность процессов, протекающих в растительном организме (фотосинтез, дыхание, водный обмен, минеральное питание, рост и развитие) и установить их взаимосвязь, чтобы научиться управлять ими.
Задачи физиологии растений. Перед современной физиологией растений стоят задачи, которые имеют большое теоретическое и практическое значение.
1. Изучение закономерностей жизнедеятельности растительного организма (механизмы питания, роста, размножения, движения)
2. Разработка теоретических основ получения максимальных урожаев на основе знаний этих механизмов).
Наряду с общими закономерностями протекание физиологических процессов, свойственных всем растениям вообще, важно знать особенности каждого вида или сорта растения и его реакцию на условия внешней среды. Важно представить и оптимальные условия произрастания того или иного вида растения. Поэтому различают наряду с общей и частные физиологии растений. В том числе и водный режим растений.
Водный режим (водообмен) растений представляет собой совокупность ряда процессов: поглощений воды растением, проведение воды по растению, потеря воды в процессе транспирации (испарение), усвоение воды клетками.
В процессе жизнедеятельности растения образуют органические вещества. Для создания одной части сухого органического вещества они расходуют 200…500 частей воды. Только за летний период культивируемые растения в среднем расходуют воды 250…700 литров на сотку. Для интенсивного роста и развития растениям требуется в два раза больше влаги, чем ее поступает с атмосферными осадками. Поэтому почвенную влагу сохраняют, растения защищают от излишнего испарения, а в отдельные периоды пополняют ее запасы. Способствует этому полив, опрыскивание, дождевание, своевременное уничтожение сорняков, рыхление и мульчирование почвы, притенение, защита растений от ветра и т. д.
Поскольку растения получают воду не только через корневую систему, но и через листья, утреннее и вечернее опрыскивание дает положительные результаты, особенно в жаркую погоду.
Большую роль в водном балансе растений играет транспирация. Она возрастает при повышении температуры воздуха, солнечной радиации, силы ветра. Когда клетки обезвоживаются, воды расходуется больше, чем ее поступает, что приводит к увяданию. В результате побеги и листья вянут, прекращаются рост растения, формирование цветковых органов, созревание плодов и т. д.
1.Общая часть.
1.1. Обзор литературы.
Корневая система является главным аппаратом поглощения воды, но не единственным: небольшое количество воды поглощается надземными частями растения во время дождей, выпадения туманов, росы. В принципе любая клетка может всасывать воду, если она не насыщена ею, даже кутикула способна разбухать.
Размеры корневых систем растений огромны. Глубина залегания корней древесных растений может достигать 20 м и более (песчаная акация, тамарикс и др.). У столетней сосны насчитывается около 5 млн. корневых окончаний, а общая длина корневой системы (без корневых волосков) достигает 50 км.
Все многообразие древесных растений по характеру строения и развития корневых систем делят на три основные группы:
1) с глубоко идущей в почву якорной корневой системой (лиственница, дуб, липа, робиния и др.);
2) с поверхностной корневой системой (ель, ясень, рябина и др.),
3) с промежуточным типом корневых систем, включающим в зависимости от лесорастительных условий как глубоко идущую, так и поверхностную корневую систему (сосна, береза, осина, многие клены, ольха и др.):
Поглощающие корни относятся к последнему порядку ветвления. Для них характерно первичное строение, отсутствие камбиального слоя. Основная их функция - поглощение воды и элементов минерального питания из почвы.
Длина всасывающей части корня травянистых растений достигает нескольких сантиметров, а древесных - всего лишь нескольких миллиметров. При этом у большинства деревьев всасывающий аппарат увеличивается за счет развития микоризы - симбиоза корня с гифами гриба. Грибной чехол микоризных корней и отходящие от него отдельные гифы существенно увеличивают поглощающую поверхность корня. Оказалось также, что микоризные растения более устойчивы к засухе, чем растения без микоризы: грибница продолжает активно всасывать воду при таком осмотическом давлении почвенного раствора, когда безмикоризные корни уже не работают.
Различают пассивное и активное поглощение воды корневой системой. Первое осуществляется за счет сосущих сил листьев, теряющих воду в процессе транспирации. Активное поглощение тесно связано с метаболизмом клеток корня и проявляется в явлениях "плача" и гуттации. "Плач" и гуттация связаны с наличием корневого давления, нагнетающего воду вверх из корней по растению. Само корневое давление является результатом жизнедеятельности корня. Движущей силой корневого давления служит энергия аэробного дыхания. Роль процесса дыхания в проявлении корневого давления может быть не только прямой, но и косвенной.
Существуют две основные гипотезы, объясняющие механизм действия корневого давления.
Осмотическая теория основана на явлениях осмоса. В сосудах центрального цилиндра поглощающего корня всегда находится раствор определенной концентрации, обладающий сосущей силой, равной целиком осмотическому давлению, так как в мертвых элементах ксилемы отсутствует тургорное давление. Поэтому вода будет отсасываться от ближайшей к сосуду клетки. Теряя воду, эта клетка будет развивать большую сосущую силу, что позволит ей отнимать воду от соседней с ней клетки и т.д. Клетки коровой паренхимы и центрального цилиндра корня будут испытывать недонасыщение водой. Этот дефицит коснется и корневого волоска, способного поглощать воду из почвы. При этом распределение величины осмотического давления в ряду клеток, проводящих воду, не играет никакой роли, ибо основным фактором осмотического поглощения воды выступает сосущая сила клеток.
Метаболическая теория, выдвинутая Д. А. Сабининым, основывается на допущении, что клетки корня, через которые наблюдается односторонний ток воды, обладают полярностью. На противоположных сторонах осуществляются диаметрально разные процессы: на одной (обращенной к периферии) синтезируются осмотически активные сахара, а на противоположной - инертный в осмотическом отношении крахмал. В первом случае осмотическое давление и сосущая сила будут больше, следовательно, к этой стороне клетки будет поступать вода от соседней клетки и выдавливаться с противоположного конца в сторону сосуда. Так создается непрерывный односторонний ток воды в корневой системе как результат развиваемого в корнях корневого давления.
Величина корневого давления обычно колеблется около 1 атм., реже 2,5...3,0 атм. С помощью корневого давления растения в ночное время суток восполняют водный дефицит, возникающий в жаркий летний день. Корневое давление у древесных растений действует в течение всего года, в том числе зимой, пополняя водные запасы ствола. Особенно велико значение корневого давления ранней весной, до распускания листьев, когда отсутствует притягивающая воду транспирация. Корневое давление можно определить с помощью манометра, прикрепленного к пеньку срезанного растения .
В обычных условиях роль пассивного поглощения воды в водообмене растений существенно выше. Оно более выгодно и в энергетическом отношении, так как не связано с затратой внутренней энергии, присущей активному поглощению, а осуществляется за счет энергии Солнца.
Путь воды в растении распадается на три различные по физиологии, строению и протяженности части:
1) По живым клеткам корня,
2) По мертвым элементам ксилемы корня, стебля, черешка и жилок,
3) По живым клеткам листа до испаряющей поверхности.
Большая часть этого пути приходится на долю водопроводящей системы, состоящей из мертвых полых сосудов у покрытосеменных и трахеид у голосеменных растений. У травянистых растений эта часть водного пути достигает нескольких или десятков сантиметров, а у древесных - многих метров.
Передвижение воды по сосудам ксилемы, длина которых может достигать нескольких десятков сантиметров, происходит довольно легко. По сосудам и трахеидам вода передвигается, как по полым трубкам, подчиняясь общим гидродинамическим законам, по живым же клеткам корня и листа - осмотическим путем, с помощью разности сосущих сил соседних клеток в правильно возрастающей последовательности. Процесс подъема воды от корней до листьев носит название восходящего тока, в отличие от нисходящего тока органических веществ от листьев к корням. Корневое давление, создающееся благодаря метаболизму корневых окончаний, получило название нижнего концевого двигателя водного тока. Притягивающие же воду сосущие силы листьев называют верхним концевым двигателем водного тока.
В зависимости от анатомического строения древесины линейная скорость восходящего тока колеблется от 1... 6 м/ч у хвойных и рассеянно-сосудистых древесных пород до 25... 60 м/ч у кольцесосудистых. Такая скорость зарегистрирована летом в полдень. Скорость передвижения воды по дереву в течение суток изменяется и в основном соответствует интенсивности транспирации.
В самом дереве быстрее всего вода передвигается в стволе и медленнее - в наиболее молодых ветвях. Среднее положение по этому показателю занимают старые ветви.
Водный ток по стволу дерева имеет следующие особенности.
С помощью изотопной техники и введения в ствол красок было показано, что у большинства древесных растений водный ток в стволе передвигается по спирали. Это тесно связано с макростроением древесного ствола, что затрудняет ответ на вопрос, какая часть корневой системы питает водой ту или иную сторону кроны дерева.
- Передвижение воды в радиальном направлении осуществляется медленнее и происходит через поры на стенках сосудов и трахеид. Оно имеет значение в поддержании нормальной оводненности живых элементов древесины и коры
- Неравномерность водного тока. Далеко не вся древесина служит местом проведения воды. У ядровых древесных растений (сосна, дуб) для этого служит только заболонь. При этом более активны в проведении воды последние годичные слои древесины. Это объясняется тем, что только эти слои переходят в древесину однолетних ветвей, тесносвязанной с водопроводящей системой листьев.
- Можно назвать и такую особенность восходящего тока в стволе дерева, как его изолированность, В общих чертах она присуща и травянистым растениям. Тем не менее, у древесных растений она выражена в большей степени. Проводящая воду древесина ствола отделена от внешней среды не только живыми клетками камбия и флоэмы, но и толстой пробкой или коркой, ограничивающих связь древесины с воздухом атмосферы.
- Такое свойство восходящего тока, как обратимость, иногда наблюдается в природе у ряда древесных пород, в том числе у некоторых хвойных (ели, пихты), при укоренении нижних ветвей, соприкасающихся с почвой, или верхушек у вываленных деревьев. Ток воды здесь идет в обратном направлении - от морфологически верхнего конца к морфо-логически нижнему концу.
Очень важна роль восходящего тока и в обеспечении живых клеток корней, ствола и ветвей кислородом, ибо проницаемость тканей коры, камбия, древесины для газов весьма низка. Этот растворенный в воде восходящего тока кислород идет на осуществление процесса дыхания живых элементов дерева.
Восходящий ток влияет и на оводненность тканей, в частности древесины ствола. Содержание воды в древесине у большинства древесных растений увеличивается от внутренней части ствола к внешней и от основания ствола к его вершине. В пределах же кроны дерева количество воды в древесине увеличивается от вершины к основанию.
У лиственных древесных пород отмечено два периода пониженной влажности - летний и во второй половине зимы и два повышенной -весенний во время сокодвижения и зимний - в первой половине зимы.
Влажность древесины молодых деревьев несколько выше, а амплитуда колебаний ее больше, чем у старых деревьев. Содержание воды в древесине меняется и в течение летних суток: наиболее высокое рано утром, а низкое - в полдень.
Вода, запасенная в середине ствола, способна передвигаться в молодые побеги, что особенно важно при отсутствии доступной для растений воды в почве (засуха, морозы). Например, 100-летняя сосна может переносить засушливый период за счет внутренних запасов воды в стволе в течение целого месяца.
Перетекающая по растению вода отличается от метаболической воды, которая непосредственно используется в различных процессах обмена веществ. Вода постоянно обменивается в клетках растений. С помощью современной техники, в частности изотопной, удалось показать, что самый быстрый обмен внутритканевой воды на внешнюю воду происходит в корнях растений, а самый медленный - в стеблях. Промежуточное положение занимают листья.
Процесс испарения воды растениями носит название транспирации. Этим термином подчеркивается отличие данного процесса от чисто физического испарения воды с поверхности какого-либо физического тела. Транспирация действительно не простое физическое испарение, а сложный физиологический процесс, который в сильной степени зависит от жизнедеятельности растения.
Физиологическая роль транспирации сводится к следующему:
-она повышает сосущую силу испаряющих клеток и создает непрерывный водный ток по растению, способствуя передвижению воды и растворенных в ней минеральных и частично органических веществ от корней к жизненно важным частям растения;
-защищает растение от перегрева прямыми солнечными лучами; температура свежего листа может быть на 7°С ниже температуры завядающего слаботранспирирующего листа;
-препятствует полному насыщению клеток водой, способствуя тем самым оптимизации целого ряда процессов метаболизма, так как при небольшом водном дефиците (до 5 %) активность синтетаз и интенсивность фотосинтеза оказываются наибольшими (В. А. Бриллиант, Н. М. Сисакян). В то же время следует обратить внимание на противоречивость функций зеленого листа: необходимость тесного контакта с атмосферой для усвоения СО2 и обеспечения листа необходимой для фотосинтеза солнечной энергией вынуждает растение развивать большую листовую поверхность. Солнечные лучи, поглощаемые хлорофиллом, нагревают листья и тем самым повышают отдачу воды в процессе транспирации. Отсюда исторически сложилась основная особенность водообмена растений, заключающаяся в перекачивании огромных количеств воды, значительно превышающих их потребности. Поэтому понятно выражение К. А. Тимирязева о том, что транспирация ассимилирующего растения является для него «неизбежным злом». Если транспирационные потери превышают поступление воды в корневые системы, то это отрицательно влияет на жизнедеятельность растений: возникает водный дефицит, завядание, остановка роста, снижение интенсивности фотосинтеза и нарушение всего метаболизма, что может привести не только к снижению продуктивности, но и к гибели растений.
Собственно испарение воды как физическое явление, т.е. переход воды из жидкого состояния в парообразное, происходит в межклетниках листа с поверхности клеток мезофилла. Образующийся пар через устьица выделяется в атмосферу. Это устьичная транспирация. В отличие от нее кутикулярная транспирация (потеря воды листом через кутикулу) характерна для молодых листьев, у которых она чуть больше 10 % по отношению к устьичной. У взрослых листьев она незначительна.
У большинства двудольных растений устьичная щель ограничена двумя замыкающими клетками полулунной формы, внутренняя стенка которых (примыкающая к щели) утолщена по сравнению со стенкой, расположенной напротив При насыщении замыкающих клеток водой и увеличении их объема наружная часть оболочки растягивается сильнее внутренней, кривизна клеток увеличивается и устьичная щель открывается .
Непременным условием открывания устьиц является развитие в замыкающих клетках достаточно высокого тургорного давления. Последнее зависит от обмена веществ и, в частности, от соотношения распада крахмала инертного в осмотическом отношении вещества, и образования осмотически активного сахара, и обратного процесса синтеза крахмала из сахара.
Наиболее употребительным показателем транспирации является ее интенсивность, под которой подразумевается количество воды, испарённой в единицу времени на единицу сырой или сухой массы или поверхности листа.
По интенсивности транспирации наиболее распространенные в лесной зоне древесные растения разделяются на три группы (табл.1):
· сильнотранспирирующие (400... 500 мг/г сырой массы в час и более);
· среднетранспирирующие (300 ...400 мг/г ч);
· лаботрансперирующие (до 200 мг/г ч
Очень важным внешним фактором, влияющим на интенсивность транспирации, является доступность воды корням растений. При недостатке воды в почве интенсивность транспирации древесных растений заметно снижается. На затопленной почве этот процесс, несмотря на обилие воды, также снижен у деревьев примерно в 1,5...2 раза. На первое место здесь выходит плохая аэрация почвы, которая вызывает ряд нежелательных изменений в метаболизме корней, а затем й надземных частей древесных растений. Снижается транспирация и при сильном охлаждении почвы в связи с резким уменьшением скорости поглощения воды. Засоление почвы также снижает транспирационные потери растений вследствие отсутствия надлежащего градиента сосущих сил между почвенным раствором и клетками поглощающих корневых окончаний.
Отсюда можно сделать следующий важный вывод: недостаток или избыток воды, засоление или холодная почва действуют на интенсивность транспирации не сами по себе, а через их влияние на поглощение воды корневыми системами.
Наряду с доступностью воды в почве значение имеют свет и влажность воздуха. На свету температура листа повышается вследствие поглощения лучей хлорофиллом, что влечет за собой и усиление водоотдачи листом. Это чисто физическое действие света. Физиологическое же воздействие света связано с известным влиянием его на движения устьиц. Показано, что даже рассеянный свет повышает интенсивность транспирации на 30.. .40 %. В темноте растения транспирируют в десятки раз слабее, чем на полном солнечном освещении.
Влажность воздуха оказывает сильное влияние на интенсивность транспирации. Согласно закону Дальтона, количество испаряемой воды прямо пропорционально дефициту насыщения воздуха водяными парами (ё), определяемому по формуле d = F - f, где F - упругость паров, насыщающих пространство при данной температуре; f - то же фактически. С увеличением разницы между F и f интенсивность транспирации, естественно, повышается. В теплицах, где влажность воздуха высокая, этот процесс протекает с низкой скоростью.
Температура воздуха действует на транспирацию прямо и косвенно. Прямое ее воздействие связано с нагреванием листа, а косвенное - через изменение упругости паров, насыщающих пространство (F). Например, если при 10 °С количество пара в воздухе достаточно для полного насыщения воздуха парами воды, то при 15 °С их будет недостаточно для этого и транспирация усилится.
Ветер способствует повышению транспирации благодаря уносу паров воды от листьев, создавая недонасыщение воздуха у их поверхности.
Различают дневной и остаточный водный дефицит. Дневной водный дефицит наблюдается в полуденные часы летнего дня. Как правило, он не вызывает существенных нарушений жизнедеятельности растений. Остаточный водный дефицит наблюдается на рассвете и свидетельствует о том, что водные запасы листа за ночь восстановились только частично вследствие низкой влажности почвы. При этом растения сначала сильно завядают, а затем при длительной засухе могут погибнуть.
Подсчёты транспирационных потерь лесом и испарением почвой и некоторые другие материалы, и учет осадков позволили определить гидрологическую роль леса в различных регионах. Оказалось, что в целом за вегетационный период лес в лесостепной зоне способствует более экономному и равномерному расходованию влаги и оказывает положительное воздействие на водный режим и урожай сельскохозяйственных культур. В таежной же зоне лес, наоборот, особенно ельники и пихтарники, расходуют воды больше, чем другая растительность остальной площади. На этом основано мелиорирующее (иссушающее) воздействие лесных экосистем.
1.2 Объект исследования и его характеристика.
Сосна Крымская. (Сосна Палласа)
Ботаническое название: Сосна Крымская.
Латинское название: Pinus pallasiana
Семейство: Сосновые (Pinaceae)
Родина: Крым, главным образом южный склон Яйлы, и Кавказ. Встречается в Малой Азии.
Освещение: светолюбива, теневынослива, жаро и засухоустойчива. Дымостойкая в городских условиях.
Почва: Предпочитает известковые почвы, но также растёт на щебневых и песчаных почвах.
Полив: нетребовательна к поливу.
Максимальная высота дерева: 20-30 (до 45м)
Средняя продолжительность жизни дерева: 500-600 лет.
Посадка: размножается семенами
Дерево: Сосна Крымская. Описание свойств и применение.


Сосна Крымская (лат. Pinus pallasiana) дерево высотой 20—30 (до 45) м. В молодости с пиромидальной, а у старых деревьев с раскидистой или зонтичной кроной. Кора нижней части ствола темно-бурая, глубоко трещиноватая, в верхней – красноватая, молодые побеги желтоватые. Хвоя по два в пучке, колючая, несколько изогнутая, длиной 8-15(18)см. темно-зеленая, очень плотная, по краю мелко-пильчатая хвоя, густо покрывает ветви. Чешуевидная хвоя светло-желтая, в отличии от темной чешуевидной хвои у сосны черная.
Ботаническое описание.
Шишки сидячие, горизонтальные, одиночные или по нескольку, коричневые, блестящие, яйцевидно-конические, длиной 5—10 см, диаметром 4,5—6 см; щитки шишечных чешуй ромбические, молодые сине-фиолетовые, зрелые буро-жёлтые, спереди широко закруглённые, выпуклые. Семена длиной 5—7 мм, тёмно-серые или почти чёрные с чёрными пятнышками; крыло длиной до 3 см, шириной 5—6 мм. В 1 кг 42 000 семян. Ветви, покрытые корой желтовато-красного цвета, широкие плоскосжатые (веерообразные), растут вертикально ориентированными и плотно прижатыми, образуют широкую конусовидную крону. Хвоя тесно прижата к ветвям, чешуевидная (у молодых, 1—2-летних растений игольчатая) с острыми верхушками, 1—3 мм длиной, светло-зелёного цвета, зимой становится бурой. Особенность хвои — отсутствие смоляных желёзок, в отличие от видов туи.
Естественный ареал.
Горы крыма до высоты 1000 метров (преимущественно на южном склоне), Северная часть Черноморского побережья Кавказа, в Малой Азии – Северная и Западная Анатолия.
Прикладное применение.
Сосна Крымская пригодна для зеленого строительства и лесомелиоративных посадок в юго-западной части прибалтики, в юго-западной Белоруссии, на Украине, а также в северном Крыму и на Северном Кавказе. Интересна как декоративное дерево для парков, а также для защитных полос и лесных насаждений, благодаря длинной темно-зеленой хвое и густому охвоению кроны весьма декоратитивна и может быть применена в садах и парках в виде солитеров и в группах; пригодна для аллей, а в крупных парках и лесопарках также в массивах, особенно при размещениях на бедных пещаных почвах, а также на каменистых склонах и меловых обнажениях. Занесена в красную книгу. В промышленном производстве даёт корабельный и строительный лес, и канифоль. В живице содержится 17—21 % скипидара; получение его промышленном значении не имеет, вследствие небольших площадей, занятых в Крыму данным видом сосны, который к тому же находятся под охраной.
Природные условия:
Воронежская обл., г. Воронеж, Дендропарк.
Подопытный экземпляр растения был получен в дендрарии ВГЛТА, г.Воронеж.
Воронежская область находится между 52˚ и 49˚ с.ш. в умеренном климатическом поясе. Лето относительно жаркое, а зима умеренно-холодная. Континентальность климата возрастает с северо-запада на юго-восток. Почти весь год территория Воронежской области находится под господством западного переноса и умеренной воздушной массы. Формирование климата происходит под влиянием умеренных, арктических и тропических воздушных масс. Среднегодовая температура воздуха составляет около 6˚ С. Средняя июльская температура — около 20,5˚С. Средняя январская температура — около −9,4˚С. Годовая амплитуда температур увеличивается к востоку области, отражая нарастания континентальности климата. Абсолютный минимум составляет −42˚С, абсолютный максимум +43˚С. Безморозный период длится от 142—157 дней на севере до 155—168 дней на юге области. Заморозки возможны во все теплые месяцы, кроме июля.
Относительная влажность воздуха заметно меняется в течение года. Так в мае-июне она составляет 41-47%, а зимой — 80-85%. Среднегодовое количество осадков меняется с северо-запада на юго-восток от 550 до 450 мм. Чуть больше осадков выпадает на наветренных склонах возвышенностей, над крупными лесными массивами, населенными пунктами. Максимум осадков приходится на июль, а минимум на февраль. В Воронежской области преобладают ветры с западной составляющей. Среднегодовая скорость ветра — 3,3-5,2 м/сек. Для зимы характерны более сильные ветры, в среднем до 6,2 м/сек.
Погодный режим Воронежской области довольно неустойчив. Зимой случаются оттепели, которые часто сменяют морозы. В зимнее время господствуют ветры западных направлений. Часто происходит вторжение воздушных масс из Западной Сибири и Арктики. Летом часто происходит смена ветра с юго-восточного на северо-западный. При этом август — самый «тихий» месяц в году со скоростью ветра от 2,7 до 4 м/сек. Лето в Воронежской области часто бывает жарким. Нередки засухи: 2-3 года на 10 лет. При засухах часты суховеи, дней с которыми бывает до 40 за теплый период. Осенью обычно заморозки начинаются в октябре, тогда же может выпасть и первый снег. Устойчивый снежный покров ложится в декабре, но бывают годы, когда он устанавливался в январе-феврале. Более того, изредка на юге области устойчивый снежный покров вообще не образуется. Весной снежный покров сходит обычно к концу марта. Происходит этот процесс очень интенсивно, занимая 2-3 недели, а в отдельные годы весь снег может растаять. Высокому уровню развития сельского хозяйства благоприятствует и набор почвенных типов. Спектр почвенных разностей Воронежской области достаточно велик - от подзолистых почв на север области до плодороднейших черноземов на юге.
1.3 Методика исследования.
Ход выполнения работы:
Для определения водообмена побега древесной породы в качестве объекта берем побег сосны Крымской.
По диаметру отверстия пробки выбираем побег и вставляем его в пробку. В опытную склянку наливаем раствор эозина, примерно на ¾ объема. Взвешиваем склянки с жидкостью (а1).
С нижней части побега взять пробу на влажность листьев. Отобрать навеску примерно 2 грамма и взвесить на тарзионных весах. Навеску поместить в бумажный пакет и указать «объект, вес, фамилию ».
Пакеты положить в сушильный шкаф при температуре 105 ˚не менее 6 часов.
После этого пакеты перенести в эксикатор для охлаждения.
Повторное взвешивание растительного материала проводить быстро, сразу после извлечения из эксикатора. Содержание воды (оводненность) рассчитывается по формуле:

а- сырая масса
б- абсолютно сухая
После этого необходимо рассчитать содержание сухого вещества:
100 – С = %
Закрепить побег в пробке таким образом чтобы нижний побег располагался на расстояние 1,5 – 2 см от дна, промежутки между побегами и пробкой замазать пластилином.
Взвесить установку с пробкой и побегом.
Оставить установку в лаборатории в обычных условиях
Экспозиция опыта 7 дней.
Через 7 дня взвесить установку с пробкой и побегом.
Извлечь пробку с побегом и взвесить склянку с оставшимся раствором эозина (м2).
1) Рассчитать количество испаренной или транспирируемой воды
Т= м1-м2
2) Рассчитать А – количество воды поглощенной побегом
А = а1-а2
3) Рассчитать транспирирующую поверхность побега исходя из того 1 гр.
Листа ____ см2
S =
Через 7 дней ветки оборвать все листья и взвесить. Посчитать транспирирующую поверхность S =
4) Определить площадь поперечного сечения древесины побега вместе с сердцевиной по формуле.
Q=3,14 * R2, см2
R- радиус древесины;
r – радиус сердцевины
5) На основании полученных данных определим интенсивность транспирации
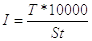
T – испаренная вода
S – поверхность хвои
t – продолжительность опыта
10000 - коэффициент перевода см2 в м2
6) Рассчитаем водный баланс
ВБ = А – Т (г)
Рассчитать скорость водного тока – количество воды прошедшей в сутки через 1 см2 поперечного сечения побега

7) Определить экономичность транспирации, какое количество воды в % от общего содержания теряется за 1 час:
Этр.= % Т – количество транспирируемой воды
% Т – количество транспирируемой воды
t – 72 часов
В – количество воды во всех листьях побега.

Д – сырая масса всех листьев
С – содержание воды в листе.
2. Специальная часть.
Количество поглощенной (А) и транспируемой воды (Т) побегом
Сосны Крымской
Таблица 1
| Объект | Поглощен. | воды, г | Испаренная | вода, г | ||
| а1 | а2 | А | m1 | m2 | Т | |
| Сосна Крымская |
Количество испаренной 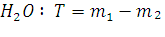
 = г
= г
 =г
=г
T = г

 = г
= г
 = г
= г
А = г
Содержание воды и сухого вещества в побегах,%
Сосны Крымской.
Таблица 2
| Объект | Оводненность хвои, % | Содержание сухого вещества, % |
| Сосна Крымская |
С = a-b/a (в %), где а - сырая масса, b - абсолютно сухая масса.
a =
b = г
C = * 100 %=г;
Содержание сухого вещества = 100% - с %
Содержание сухого вещества = %
Транспирирующая поверхность побега, Сосны Крымской.
Таблица 3
| Объект | Вес хвои, г. | Площадь хвои (S), |
| Сосна Крымская |
Д - масса всей хвои
Д = г
Площадь хвои:
1г =  ; -х
; -х
х = * =
Интенсивность транспирации, Сосна Крымская, 
Таблица 4
| Объект | Транспирируем. вода (Т), г | Поверхность хвои, | Продолжительность опыта, часы | Интенсивн. транспирации г/м ч |
| Сосна Крымская |

 ,
, 
Т =
S =
t = ч
ИТ = г/м
Водный баланс Сосны Крымской, г
Таблица 5
| Объект | Поглощ. вода (А), г | Испареню вода (Т), г | Водный баланс, г |
| Сосна Крымская |
Водный баланс = А-Т
А = г
Т = г
Водный баланс = г
Таблица 6
Скорость водного тока (V) Сосны Крымской, 
| Объект | Поглощенная вода, г,А | Испаренная вода, г,Т | Среднее из поглощ. и испаренной воды, г A+T/2 | Поперечное сечение дрвесины, | Скорость водного тока, V |
| Сосна Крымская |
 Q
Q
V = M/Q
V - скорость водного тока
М - среднее из массы поглощенной и испаренной воды, г
М = (А+Т)/2
М = г
Площадь поперечного сечения древесины Q = 
R = мм
Q = мм
V = 
Экономность транспирации Сосны Крымской, % ( )
)
Таблица7
| Объект | Транспир. вода Т, г | Продолжительность опыта, t (часы) | Масса всей хвои, г, Д | % в хвое, С | Кол. Воды в хвое |
% |
| Сосна Крымская | 2 |

 = 100×T/t×В
= 100×T/t×В
Т = г
t = ч
B = Д × с/100, %
В = %
= %
Выводы:
1. По данным - содержание воды, сухого вещества составило
%,
2. По данным определить сколько раз вода в побеге заменялась в
течение опыта ___раз.
3. Объяснить, почему водный баланс не может быть положительным
или отрицательным.
4. Скорость водного тока .
5. ИТ хвои г/м ч.
Список используемой литературы.
1.Веретенников «Физиология растений», Воронеж, ВГЛТА,2002.
2.http://ru.wikipedia.org/
3.http://dic.academic.ru/dic.nsf/enc_geo/6982/природные условия.
4.Кузнецов, В.В. «Физиология растений»,- М.: Абрис, 2011.
 (zip - application/zip)
(zip - application/zip)